Revise generic
boundaries in the Buteo group
Proposal (460) to South American
Classification Committee
Effect on South American CL: This would revise generic boundaries extensively in Buteo and Leucopternis.
Background
& New Information: For
several years, we’ve had plenty of indication that the current boundaries of
the genera Buteo, Leucopternis, and relatives in our
current classification are a mess.
Raposo do Amaral et al. (2009) have produced a comprehensive phylogeny
of buteonine hawks, and their data will form the primary basis for this
proposal. Findings from earlier
papers (see Notes below) are largely consistent with Raposo do Amaral et al.
(2009) and will not be discussed further.
Two of the relevant Notes from our SACC classification are:
12a. Genetic data (Raposo et al. 2006, Lerner et al. 2008, Raposo do
Amaral et al. 2009) indicate that the genus Leucopternis is
polyphyletic; as currently defined, Leucopternis includes at least three
distinct groups that are not each others' closest relatives: (1) L. melanops,
L. kuhli, and L. semiplumbeus; (2) L. albicollis,
L. occidentalis, and L. polionotus; (3) L. plumbeus,
L. schistaceus, L. lacernulatus, which are intermingled within a
group with Buteogallus and Harpyhaliaetus; and (4) L. princeps, whose placement is uncertain. Raposo do Amaral et al. (2009)
recommended placing princeps in a
monotypic genus Morphnarchus, plumbeus in a new monotypic genus Cryptoleucopteryx, schistaceus in Buteogallus,
and lacernulatus in a new monotypic
genus Amadonastur. SACC proposals
needed.
18. Genetic data (Riesing et al. 2003) indicate that Geranoaetus is the
sister taxon to Buteo polyosoma/B. poecilochrous
and that maintenance of a monotypic genus is not warranted; it had been placed
in Buteo formerly (e.g., Wetmore 1933, Hellmayr & Conover 1949,
Friedmann 1950), but recent authors have generally followed Amadon (1963), who
suggested that it might be closer to Buteogallus or Leucopternis
than to Buteo. Clark (2006) disputed Amadon's rationale for maintaining
it is a genus separate from Buteo. SACC proposal to merge Geranoaetus
into Buteo did not pass. New genetic data (Lerner et al. 2008)
provide even stronger evidence for merger of Geranoaetus, at least as
currently defined, because it is the sister species to B. polyosoma. SACC proposal to merge Geranoaetus
into Buteo did not pass. Raposo do Amaral et al. (2009) further
confirmed that Geranoaetus is
the sister to Buteo polyosoma sensu lato. SACC Proposal
needed.
Raposo do
Amaral et al.’s (2009) taxon sampling (105 specimens, 54 species) and gene
sampling (6000 bp of 9 genes, mitochondrial and nuclear) is exemplary. I doubt that anyone will produce a
better data set anytime soon. This
proposal deals only with their Group H, whose monophyly has excellent support;
the relevant portion of their tree (from their Fig. 3) is pasted in here (sorry
for the poor resolution of the screen grab):
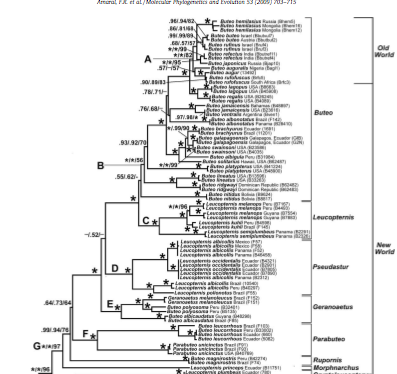
The Buteo group itself (Group G) is strongly
supported as a monophyletic group as is sister relationship to the Buteogallus group (Group H). It includes everything in our current
classification in Buteo plus Parabuteo, Geranoaetus, and most Leucopternis
(minus the 4 species that are part of Group H; see Proposal 459).
Analysis
and Recommendation: Virtually every critical node in Group G’s tree has
strong support. Our current Buteo and Leucopternis are both polyphyletic, and so changes must be
made. So, the only point of real
discussion is the subjective exercise of how broadly to delimit the
genera. Raposo do Amaral et al.
have defined these very narrowly.
However, one option would be to expand Buteo to include all species in Group G. Even the two outliers, “Leucopternis”
princeps (placed by Raposo do Amaral
et al. in a resurrected monotypic genus Morphnarchus)
and Buteo magnirostris (placed by Raposo do Amaral et al. in a resurrected
monotypic genus Rupornis), don’t
really stand out (to me anyway) as beyond the range of variation encompassed
even with a narrow Buteo). In my subjective view, there is so much
variation among the species in narrowly defined Buteo in terms of color, shape, behavior, and size that adding the
other 6 genera recognized by Raposo do Amaral et al. does not really add to its
phenotypic heterogeneity. Narrow Buteo includes heavy, robust species of
open country such as B. regalis and
small chunky species of forest such as B.
platypterus. It includes B. nitidus, long placed in a monotypic
genus Asturina. It includes blackish species such as B. albonotatus,
gray-plumaged species such as B. nitidus,
and relatively pale species (ventrally) such as B. brachyurus. It also
includes all Old World Buteo sampled
so far. More importantly, Raposo
do Amaral’s et al.’s Buteo is not
unequivocally monophyletic if nitidus
is included, and in fact one could make a case for resurrection of Asturina based on weak support for the
node that links it to core Buteo.
A YES vote
on this proposal would be for adopting Raposo do Amaral et al. as is, namely
recognize 7 genera (in the following sequence) for the group instead of 4, as
follows:
Morphnarchus
(monotypic; formerly L. princeps)
Rupornis
(monotypic; formerly B. magnirostris)
Parabuteo (unicinctus plus leucorrhous)
Geranoaetus (to also
include B. polyosoma and B. albicaudatus, a combination we’ve
voted down previously)
Pseudastur (resurrected
for L. albicollis/occidentalis; this might actually become
monotypic given that albicollis is
evidently paraphyletic with respect to occidentalis
in their tree)
Leucopternis (melanops, kuhli, semiplumbeus)
Buteo (nitidus, platypterus, albigula, brachyurus,
swainsoni, galapagoensis, albonotatus, ventralis)
A NO vote
would be to broaden generic boundaries, from small tweaks to as much as
including everything in Buteo. If this proposal fails, I’ll write
additional proposals to take into account broader generic limits. I do not have a strong recommendation,
but I am going to vote NO, given the points made about on the heterogeneity of
narrow Buteo. Because delimiting genera is a
subjective exercise as long as each is monophyletic, I hope to hear others
opinions, and my NO vote is not a firm one.
Literature Cited:
RAPOSO
DO AMARAL, F., F. H. SHELDON, A. GAMAUF, E. HARING, M. RIESING, L. F. SILVEIRA,
AND A. WAJNTAL. 2009. Patterns and processes of
diversification in a widespread and ecologically diverse avian group, the
buteonine hawks (Aves, Accipitridae). Molecular Phylogenetics and Evolution 53:
703-715.
Van Remsen, August 2010
=========================================================
Comments from Bret Whitney: “I like Rupornis for Roadside Hawk. It’s a bizarre bird, frankly, a highly
successful New World raptor not quite like anything else, intermediate
morphologically and behaviorally between Accipiter
and small Buteo with distinctive
vocalization patterns and behaviors (nothing else is even close to as
consistently vocal as Rupornis, for
instance). To lump it into Buteo just obscures the picture, in my
opinion. Yeah, I know, having such
different types as Ferrug and Broad-winged in one genus is also “uncomfortable”
-- but I guess the best path forward is to split out only what seems clearly
divergent when ornithologists can agree that there’s good concordance among
various datasets.”
Comments from Robbins: “NO, to be consistent with my comments under
proposal 459. I would also add
vocalizations to Van’s comments about variation in Buteo; it is considerable among even closely allied species.”
Comments
solicited from Fabio Raposo:
“Thanks very much to Van and
the committee for requesting comments and letting us be part of this
discussion. A few important points
on this one (for more general comments on the buteonine proposals, please see
SACC #459):
Yes, there is a lot of
heterogeneity in Buteo, even when narrowly defined as we did. However, the other monophyletic groups
treated in our paper as genera are far from heterogeneous, since they have
clear identities as groups (see below, we explicitly indicate why we
strongly support that point of view). Exactly for this reason, we don’t
understand why an enormous, uninformative and undiagnosable Buteo
would be a better option - why add more heterogeneity to Buteo, when we
can indicate specific characteristics of each of those
clades? Furthermore, our classification is "testable" - those
with access to a good bird collection will clearly see those groups/characters
in 10 minutes with the birds on a table (this is something that we did over and
over again before closing this paper), while a catchall Buteo has no
concrete identity as a group. A huge, undiagnosable, species-hungry Buteo
(by the way, rejected by the committee twice when dealing with Geranoaetus
- see SACC #282 and SACC #387) would be unnecessarily in the top 10
largest avian genera ever.
--
genus Leucopternis (L.
semiplumbeus, L. kuhli and L. melanops): generally speaking,
small, black and white Neotropical buteonine hawks, found exclusively in
forests. Plain white underparts, narrow black streaks or solid gray on the
sides of the neck; short wings (primaries extending to less than half of tail);
plain black tails with one medial white band in adults (and two white bands in
young birds).
--
genus Pseudastur (P.
albicollis, P. occidentalis and P. polionotus): generally
speaking, large, black and white Neotropical buteonine hawks - found
exclusively in forests. White underparts, plain white tails with a single
black band, and secondaries and tertials with conspicuous white tips.
-- genus Geranoaetus (G. melanoleucus,
G. polyosoma and G. albicaudatus): generally speaking, large,
dark headed open vegetation buteonine hawks - mostly Neotropical. Densely
feathered faces, solid black or gray sides of the neck, white underparts with
flanks (and sometimes belly) finely streaked, long primaries that reach the tip
of the tail; densely barred tails with narrow blackish bars and a subterminal
large black band, or entirely solid black.
-- genus Parabuteo (P. unicinctus and
P. leucorrhous): Plumage is strikingly similar between those two
species - overall black or dark brown coloration, white rump and undertail
coverts, and mostly rufous or light brown thighs.
On the other hand, Rupornis magnirostris
(as well indicated by Bret) and Morphnarchus princeps are
very divergent species, isolated from the rest of the tree by lots of
evolutionary time and autapomorphic characteristics - and that’s why monotypic
genera fit best for those two.
Finally, a correction to the
proposal: Pseudastur also includes L. polionotus, which is
clearly a good species, sister to the Amazonian subspecies of L. albicollis.
So even if the trans-Andean L. albicollis subspecies are lumped
with L. occidentalis, there still would be three species (L.
polionotus, L. albicollis from Amazonia, and trans-Andean L.
albicollis+L. occidentalis) - so nothing to worry about this genus
becoming monotypic.
Comments from Zimmer: “YES.
I tried to be open to the idea of throwing everything into a broadly
defined Buteo, but I just can’t do
it. I think Raposo do Amaral et
al. have actually hit on a pretty sound classification scheme that makes a lot
of sense with respect to generic limits, and which is supported by molecular
data. In looking at these
groupings, certain things jump out at me.
For starters, I think L. princeps
is an odd beast that doesn’t clearly align with anything else. It is very vocal, and prone to
delivering these vocalizations while soaring high above the forest canopy,
almost in the manner of Spizaetus
tyrannus. Bret has already
commented on the oddity of magnirostris,
and how exceptionally vocal it is, and how it is almost intermediate between Buteo and Accipiter. Then you
have Parabuteo. I’m not sure how good of a fit leucorrhous and unicinctus are with one another, but in looking at unicinctus, you’re dealing with a
group-hunting, polyandrous or cooperative-breeding species whose social system
and ecology would seem to place it somewhere different than just about
everything else {Except for galapagoensis;
but that seems like a different case, given that it is a rare resident of small
equatorial islands, operating in an environment of competitive release and
potential saturation of available territories}. Pseudastur and Leucopternis (as redefined by Raposo do
Amaral et al.) seem like natural groupings on vocal, morphological and
ecological grounds. The remaining Buteo species seem fine. As Van notes, you could make a case for
resurrection of Asturina for nitidus, although in many of its quirks
it reminds me of Buteo lineatus. All in all, there are some things here
I could quibble with (most notably, the linkage of unicinctus and leucorrhous),
but I think this proposed classification has a lot of advantages over
recognizing a single, broadly defined Buteo. As Van and Mark point out, even
narrowly defined Buteo is very
heterogeneous on morphological and vocal grounds, but throwing all of these
birds together would result in much greater behavioral/ecological heterogeneity
(including patterns of vocal behavior as opposed to just looking at comparative
differences in voices) than what I currently see in Buteo.”
Comments from Stiles: “YES – each of the groups seems reasonably coherent, and I would say diagnosable. Unlike Van I definitely feel that magnirostris is an oddball in Buteo: it is much more vocal than any other, and is the only one that I would call a poor soarer; it is rather more lightly built as well; I am not at all averse to separating it in Rupornis. The rest of the buteos vary considerably in size and color but morphologically seem to me to be a relatively homogeneous assemblage, and the phylogeny doesn’t facilitate division into more groups (I regard nitidus as a relatively typical Buteo, much more so than magnirostris, and have never been impressed with separating it in Asturina). As for Geranoaetus, this also seems to me a rather coherent group sharing complex plumage sequences and a fairly common color scheme; whether SACC recognizes it as a species or not, morphologically poecilochrous is intermediate between polyosoma-albicaudatus and melanoleucus in its broader wings and shorter tail. The division of Leucopternis into two groups is mandated by the phylogeny and I have always considered princeps to be “special” – its aerial acrobatics and loud, melodious flight “song” bring to mind Spizaetus tyrannus more than any other Leucopternis or Buteo, whereas its stolid demeanor when perched and hunting tactics remind me more of Buteogallus: here too, I am not upset by placing it in its own genus. The inclusion of leucorrhous in Parabuteo makes sense in terms of color and pattern. All told, I think that the Raposo et al. phylogeny is a major step forward and I am in agreement with their generic allocations.”
Comments from Nores: “YES. A pesar de que Remsen
tiene razón en que hay una gran variabilidad en el género Buteo, incluso
en un angostamente definido Buteo, no me conforma de manera alguna
poner todas las especies dentro de un Buteo expandido. Particularmente
me agrada la idea de separar Buteo magnirostris de los otros Buteo,
porque siempre me pareció muy diferente, tanto en aspecto como en
comportamiento.”
Comments solicited from Bill Clark: “I am completely in favor of Proposal 460 to separate the buteonines in the genera Buteo and Leucopternis of South America into multiple genera. The grouping proposed by Raposo do Amaral et al. makes a lot of sense to me. The English name for the South American species nitidus, should now be called now ‘Gray-lined Hawk’, based on the paper by Brian Millsap, Sergio Seipke, and I that will advocate splitting the two gray hawks; it had been accepted by Condor. We three agree that both forms belong in Buteo, not separated in Asturina. Plumbeous Hawk occurs in South America along the Colombian coast and should be included in the genus Leucopternis. Regarding the comments of Zimmer on unicinctus, there are no reports, to my knowledge, of the South American form group hunting or cooperative breeding; does anyone know of such? (Sergio Seipke and I will be examining the differences between the SA and northern forms of this taxon in the near future.)”
Comments from Pacheco: “YES. Considero o arranjo genérico proposto pelo trabalho de Fabio Raposo e colegas um avanço real ao entendimento geral das interrelações deste grupo. Ademais, como já dito pelo colegas, ele reforça o senso comum e coloca os “esquisitos” L. princeps e Roadside Hawk em posição privativa.”
Comments solicited from David Mindell: “For me and probably most of you, genus assignments are inevitably
subjective (hence the voting), excepting the valuable criterion of monophyly. I
have looked at a range of specimens for all the taxa and have seen most though
not all in the field. But, there is no standard or metric regarding degree of
difference in morphology, physiology, life history, behavior or niche breadth,
that can be broadly and objectively applied to guide taxonomic category
delineation across disparate groups in general and Accipitridae in particular.
I think the only/best objective criterion for assignment of genus
boundaries is time. Time is a physical constant and metric reliably quantified
by radiometric methods (assuming provision of reliable confidence
intervals). Time passes at a regular and independent rate in comparison
to rates of change in organismal features, and there are no subjective
disagreements about the relative importance of this or that millenium as there
are for traits enabling this or that form of feeding, metabolism, communication
or even replication. Further, passage of time can be measured and
recognized, throughout life's history and across the tree of life topology,
whereas the histories of particular organismal or avian traits, that might
inform the ranking of categories, are less easily discovered, due to
convergence, selection and chance events among other things.
“So, you won't be
surprised that my 'vote' on the proposal would be 'no'. It wouldn't
bother me to have an unusually large genus Buteo (among
the largest 10-20 for Aves as one commentator notes), particularly if this were
supported by recent species divergence time estimates comparable to those for
other speciose genera. It would be
nice to be able to compare ranked categories across avian families or orders. One might learn more about evolutionary
history, and communicate it better through classification if an objective
criterion were applied. But that's a digression...
“In any event, it is
nice to see and participate in improving phylogenetic resolution for this
group, with remarkably good congruence among datasets and research teams.
I do see the real value of binomials for the public and resource managers
(and even evolutionary biologists) among others, so am not dissing the topic at
all.”
Comments from Jaramillo: “YES. This makes a great deal of sense to me, even the broader Geranoaetus. Immature plumages of polyosoma and the sequence of maturation actually mirrors much of what is seen in Geranoaetus melanoleucus; furthermore juvenile melanoleucus has a reasonably standard tail size clarifying that the short adult tail is a derived feature only present in one age group of the species. A note about Parabuteo unicinctus, in the far south (Chile – Argentina) this species does not appear to be social, and ecologically is quite different from the desert-dwelling population in Mexico – USA etc. So that unique character varies within the species.”