Proposal (469) to South American Classification Committee
Species
limits in Paroaria
Effect on South
American CL: this proposal would elevate to
species rank various forms within Paroaria
that are currently treated as subspecies.
Background: Species limits in the genus Paroaria have always been somewhat arbitrary. Members of this genus are all adorned by red
somewhere on the face or throat, and variably developed crest from none to well
developed. The members differ in
distribution or lack of black on the face, bill coloration and upperparts
coloration (grayish to blackish). All
are whitish below. Here’s what our
current footnote states:
66. Paroaria gularis, P. baeri, and P. capitata
form a superspecies (Sibley & Monroe 1990); evidence for treating them as
separate species is weak (Paynter 1970a); Hellmayr (1938) suspected that P.
capitata might best be treated as a subspecies of P. gularis,
and Meyer de Schauensee (1966) suspected that baeri might also
best be treated as a subspecies of P. gularis. The subspecies P. g. nigrogenis of Venezuela was formerly (e.g., REF)
treated as a separate species from Paroaria gularis. Dávalos & Porzecanski (2009) also found
evidence that nigrogenis was not most
closely to P. gularis; they elevated
all diagnosable taxa to species rank based on the phylogenetic species concept.
New information: Dávalos and
Porzecanski (2009) published a paper based on molecular as well as a
morphological analysis of the genus. They examined over 600 specimens measured
and scored them for plumage characters.
Molecular data included complete sequences of cytochrome b. Note that they did not have molecular data
for xinguensis.
They characterized species based on
morphological data, in particular whether characters shared by individuals were
of a unique combination and restricted to a geographic region. When these unique characters were fixed in
all individuals sampled, the population was considered diagnosably distinct and
a hypothesized phylogenetic species. The
molecular data was then looked at with at least two individuals per
hypothesized phylogenetic species looking for unique fixed mutations for
support of these as valid species.
They suggest separating the genus into
8 species based on these data:
coronata - 24
unique substitutions in the cytochrome b gene
dominicana - 33 unique substitutions in the cytochrome b gene
nigrogenis - 20 unique
substitutions in the cytochrome b gene
xinguensis – no
molecular data.
baeri - 11
unique substitutions in the cytochrome b gene, but since sequences of
its sister xinguensis, are lacking, this may be an overestimate.
gularis - two unique substitutions in the cytochrome b gene
cervicalis - two unique substitutions in the cytochrome b gene
capitata - five
unique substitutions in the cytochrome b gene
They stated the following: “Lumping of nigrogenis, cervicalis
and gularis is not supported by
genetic or morphological data.” This is perhaps the most clear-cut result in
this study. The taxon nigrogenis does not appear to be closely
related to gularis; it is well
differentiated based on the molecular data, and well differentiated based on
the morphological data. Note also that
Restall et al. (2006) separated nigrogenis
from gularis and note that the two
are sympatric in extreme SW Venezuela with no evidence of hybridization. These data strongly suggest that nigrogenis is a valid biological
species.
The relationship of capitata,
cervicalis, and gularis is more problematic. All share a similar color pattern with
red head, black throat and breast. The
species capitata has an entirely
yellow bill, unlike cervicalis and gularis.
The form cervicalis has no
black around the eyes, whereas gularis
does have a small dark area around the eyes; this is essentially the main
difference between the two and does not seem to be all that impressive to
me. Vocal data would be good to have to
compare. Furthermore, the type of cervicalis is apparently aberrant,
further muddying this situation. The
apparent fixed genetic differences are of interest, although only two cervicalis were sampled, a larger sample
size would have been better. The range
of cervicalis and gularis likely meet somewhere in W
Brazil or NE Bolivia and sympatry should be looked for, and any evidence of
hybridization. The plot of ML distances
showed the gularis-cervicalis comparison to be intermediate
between within-species variation and between-species variation, in other words
more grey than black or white. At this
point it does not seem clear to me that cervicalis
warrants separation as a biological species.
On the other hand, capitata is
geographically isolated from cervicalis-gularis and shows a greater degree of
differentiation morphologically and molecularly. Retaining it as separate seems
reasonable.
Similarly, the division of xinguensis from baeri is
weak. No molecular data were available for xinguensis
to begin with. The form is diagnosable in that it has a black throat instead of
the red throat of baeri, yet only 5
specimens in total for both of these taxa were studied. It seems very premature
to separate xinguensis from baeri at this point.
I think that it is best to separate the
issues into two parts:
A. Elevate nigrogenis to species rank.
Masked Cardinal (used in Restall et al 2006) is an appropriate English
name given that it is the only one with a bold black stripe through the face in
this genus of “cardinals.”
B. Also elevate the following to
species rank: cervicalis and xinguensis.
Recommendation:
I recommend a YES on A and a NO on
B. Dávalos and Porzecanski regard all
diagnosable units as species, using PSC, so there is really no evidence for any
of the other diagnosable taxa being ranked as species under BSC.
Literature Cited.
Dávalos, L.M., Porzecanski, A.L. (2009)
Accounting for molecular stochasticity in systematic revisions: species limits
and phylogeny of Paroaria. Molecular
Phylogenetics and Evolution 53:
234-248.
RESTALL, R.,
C. RODNER, AND M. LENTINO. 2006. Birds of northern South America. An
identification guide. Christopher Helm, London.
Alvaro Jaramillo, October 2010
=========================================================
Comments from Stiles: A.
YES – both gularis and nigrogenis also occur in NE Colombia at
no great distance from one another; although I am not aware of any locality
where both have been taken here, I can also detect no indication of
intermediacy in any of the specimens here. B. NO – I agree with Alvaro that
evidence for splitting these two is at present inconclusive.”
Comments from Nores: “A.
YES. Aunque por distribución parece corresponder a una subespecie, los
datos moleculares y el análisis hecho en la propuesta por Álvaro indican que se
trata de una especie. B. NO. Coincido con Álvaro que no hay
evidencias concluyentes como para elevar estas subespecies al rango de especies.”
Comments from Robbins: “A. YES.
Both molecular and morphological data support this conclusion. B. NO, until additional data are presented
supporting elevating these to species status.”
Comments from Remsen: A. YES.
But only because of Restall et al.’s report of sympatry, of which I was
not aware, and Gary’s report of near parapatry.
Otherwise, I see no evidence in this paper, either morphological or
genetic, that supports species rank for nigrogenis. That nigrogularis
does not group with gularis might be
a simple gene tree vs. species tree problem, given that only one mtDNA was
sequenced. B. NO. The authors of the paper have documented that
there are 8 diagnosable taxa based on plumage.
Whether those diagnosable units are ranked as valid subspecies or PSC
species is a matter of taste - - the results are still the same. Any use of comparative genetic distances for
a single locus for assigning taxon rank is naïve. That there are fixed genetic differences in
cyt b among taxa known to also have diagnosable, presumably genetically based
plumage differences is of interest but how that relates to taxon rank is a
mystery to me. If there were no fixed
differences in cyt b, but fixed plumage differences, would the authors have
arrived at a separate taxonomic decision?”
Comments from Zimmer: “A. YES, based on reports of sympatry and parapatry with gularis, coupled with diagnosable plumage differences that are on a scale with some of the differences between currently recognized species in the genus. B. NO. I see nothing in the paper by Dávalos and Porzecanski to indicate that cervicalis and xinguensis are anything more than diagnosable subspecies/phylogenetic species, which is how we currently treat them.”
Comments from Pacheco: “A.
Yes. A julgar pela aparente
ausência de espécimes intermediários entre os táxons nigrogenis e gularis, no sw.
Venezuela e, possivelmente, no nw Colômbia – sou favorável ao status de espécie
para nigrogenis sob o BSC. B. No. Em vista das informações disponíveis é
prematuro considerar cervicalis e, sobretudo, xinguensis como
espécies plenas sob o BSC.”
Comments
from Perez: “Paroaria
gularis and nigrogenis
are largely divergent in both molecular and morphological characters; however,
the key data supporting species rank was the following information:
“Note
also that Restall et al. (2006) separated nigrogenis from gularis
and note that the two are sympatric in extreme SW Venezuela with no evidence of
hybridization. These data strongly suggest that nigrogenis is a
valid biological species.”
“Although Gary commented on another
potential zone of potential parapatry without records of intermediate
specimens, the SW Venezuelan zone of sympatry was the key issue.
“Following this decision, I was
interested in doing a more in-depth research in the area to describe this zone
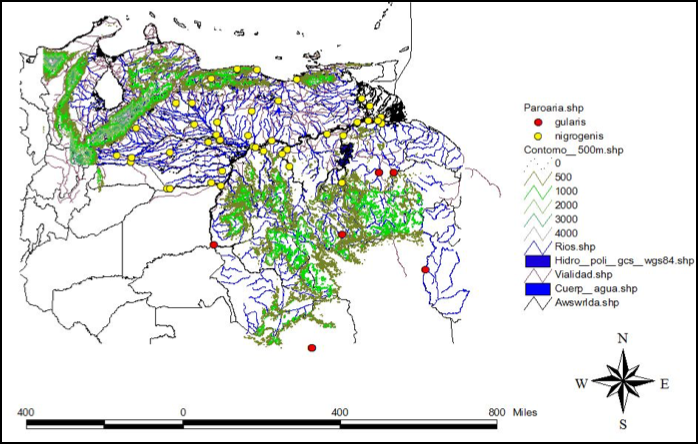
of sympatry or find a potential area for a contact zone study between both gularis and nigrogenis. Together with Miguel Lentino, from the Colección
Ornitológica Phelps (COP), we produced a map (see below) including all records
of these species present in three of the most important ornithological
collections in Venezuela (with specimens from SW Venezuela). To our surprise,
we did not find any confirmation of sympatry; in fact, the closest records
between both taxa were about 200 km from each other. We also requested
information for many birdwatchers, who routinely travel around the area, and
found no evidence for such claimed sympatry. We did find, however, two
misidentified COP specimens collected at El Dorado, at SW Venezuela. Those
specimens were labeled as nigrogenis
whereas the rest of specimens in the same locality were gularis, potentially misleading description of biogeographic patterns.
The question now is if we will keep our decision to elevate nigrogenis to species rank, based only
on the potential parapatry in Colombia, and the large morphological (even in
juveniles) and molecular differentiation between gularis and nigrogenis. At
least, I think we need to discuss if such differences are enough to claim
reproductive isolation under the biological species concept we use here (which
I think will be difficult); if not, we might need to reconsider our decision.”
New Comment from Remsen: “Jorge’s comments
and research leads me to change my vote to NO.
The primary reason I voted for this was the alleged sympatry (and given
what I perceive as limited meaningful differences among these parapatric taxa,
I was always suspicious).”