Proposal (555) to South
American Classification Committee
Reclassification
of the Scolopacidae
Effect
on SACC: This proposal would divide our current family
Scolopacidae into 5 subfamilies and change the sequence of genera within the
family.
Background:
Our current classification does not include subfamily structure and
follows a traditional sequence of genera.
The reason for the lack of subfamily structure was our wise assessment
of the lack of concordance among other classifications and the absence of a
solid DNA-based phylogeny. Our footnote
is as follows:
1. <note on genera, linear sequence> Jehl (1968b). The
family Scolopacidae is traditionally split into five or more subfamilies and
additional tribes (e.g., AOU 1998). Livezey (2010) recognized four
subfamilies (Arenariinae, Calidrinae, Tringinae, Scolopacinae) and maintained
the phalaropes as a separate family. Genetic data
(e.g. Gibson & Baker 2012), however, provide very weak or no support for
the monophyly of these groups, and although the phalaropes are monophyletic,
they are deeply embedded in the Scolopacidae and sister to the tringines. Gibson & Baker (2012) identified five
major lineages in the family. SACC proposal needed to recognize five subfamilies.
New information: Gibson & Baker (2012) produced a
phylogenetic hypothesis based on strong taxon-sampling (84 species) and good
gene-sampling (6300+ bps; 1 nuclear + 4 mitochondrial genes). Branching patterns in general were well-resolved
(except for problems within the explosive calidridine radiation). Here is their tree – sorry about the poor
resolution – if anyone needs a pdf, just let me know:
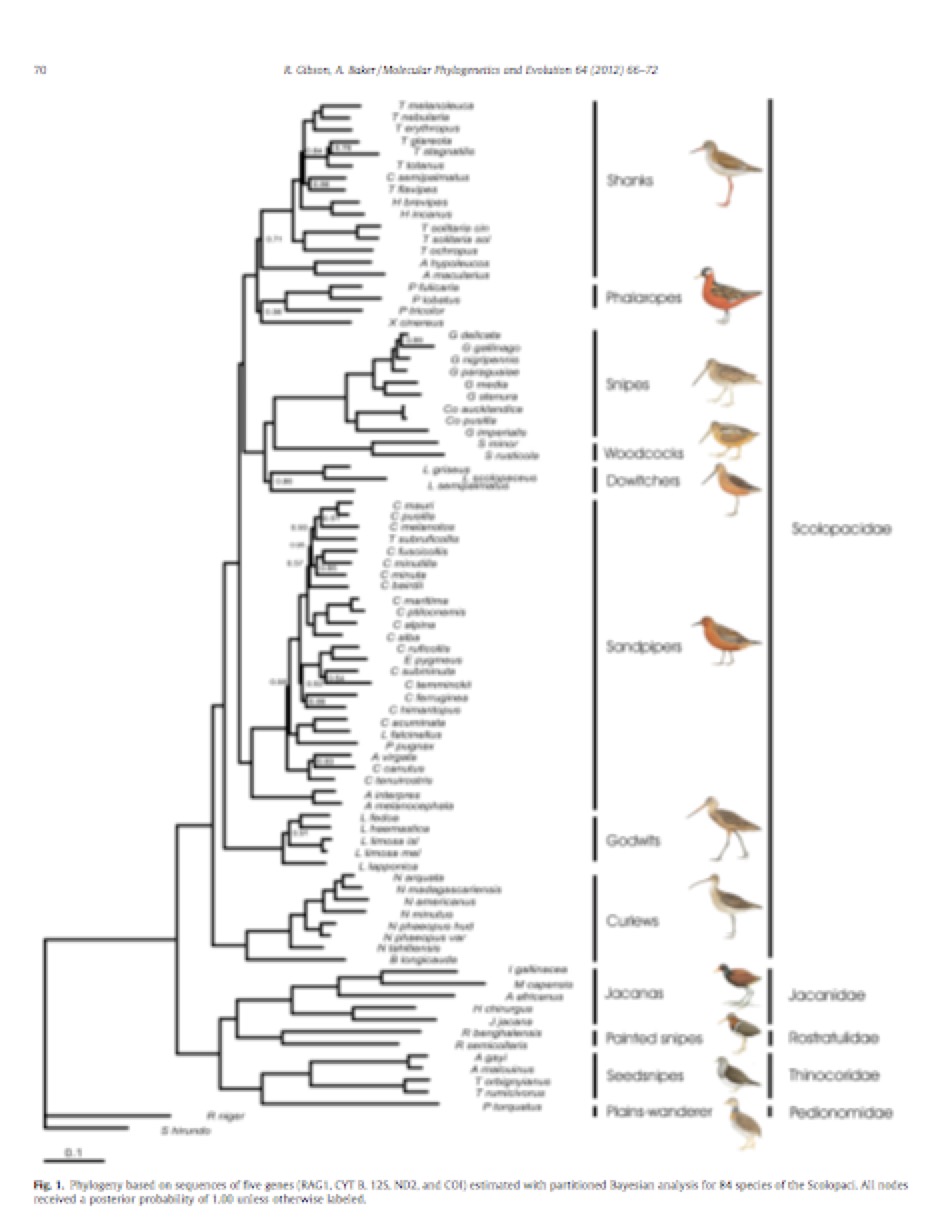
I
suggest that the best interpretation of the tree is to recognize 5 major
lineages and to give each one subfamily rank.
Note that Gibson & Baker name 8 groups, not 5, which would be an
alternative treatment, i.e. 8 subfamilies.
I suggest condensation to five for the following reasons.
(1) The
Snipe-Dowitcher-Woodcock group is probably best treated as a single group
because the branch lengths connecting them are very short, inclusion of Limnodromus semipalmatus in the
Dowitchers has weak support, and taxon-sampling within the Woodcock and Snipe
groups is weak (including absence of Limnocryptes).
(2) The
Phalaropes if treated as a subfamily would technically also have to include Xenus (Terek Sandpiper), which groups
with them albeit with substandard support.
That result is best treated as anomalous, in my opinion, until
corroborated by additional data (parallel to the initial anomalous results for Pluvialis; see Baker et al. 2012). An alternative approach would be to recognize
the Phalaropes as a subfamily and to retain Xenus
in its traditional spot in the Tringinae or place it as Incertae Sedis. Although not giving the phalaropes subfamily
rank might seem extreme, keep in mind that Phalaropus
tricolor (which really deserves to have its monotypic genus status
reinstated: Steganopus) strikes many
field people as having some tringine-like behaviors.
Tangentially,
I take the opportunity to point out that Livezey (2010) placed the phalaropes
in their own family, as they were for most of their history due to their
unusual morphology. Assuming that the
gene-based phylogeny reflects historical relationships, this is yet another
warning for use of morphological data to infer phylogeny. Livezey’s careful and quantitative analysis
of hundreds of phenotypic characters was unable to distinguish highly derived
morphological change from lineage history, just as it was unable to identify
strongly convergent morphology (e.g., Livezey’s sister relationship between
Podicipedidae and Gaviidae).
Another
point worth considering is giving subfamily rank to the Turnstones. Although not labeled as a group by Gibson
& Baker, just looking at relative divergence, they deserve naming if
Woodcocks, Snipes, and Dowitchers do, and their distinctive morphology has led
to their classification as a separate family or subfamily in the past. But looking at branch lengths and node depths
in the tree, if turnstones are treated as a subfamily, then snipes, woodcocks,
and dowitchers also should be. As
predicted from any categorical scheme inflicted on a continuum of
differentiation, borderline situations are inevitable. I suggest that that if we decide to include
the Tribe level in our classification, then turnstones would be a prime
candidate for recognition at that level.
Taking
the Gibson-Baker tree and transforming it to a classification for species in
the SACC area, including changes in species sequence to match the tree, would
look like this:
SCOLOPACIDAE
(SANDPIPERS)
Numeniinae
Bartramia longicauda Upland Sandpiper
Numenius borealis Eskimo Curlew
Numenius phaeopus Whimbrel
Numenius americanus Long-billed Curlew
Limosinae
Limosa
lapponica Bar-tailed Godwit
Limosa limosa Black-tailed Godwit
Limosa haemastica Hudsonian Godwit
Limosa fedoa Marbled Godwit
Arenariinae
Arenaria
interpres Ruddy Turnstone
Arenaria melanocephala Black Turnstone
Aphriza virgata Surfbird
Calidris canutus Red Knot
Calidris alba Sanderling
Calidris pusilla Semipalmated Sandpiper
Calidris mauri Western Sandpiper
Calidris minutilla Least Sandpiper
Calidris fuscicollis White-rumped Sandpiper
Calidris bairdii Baird's Sandpiper
Calidris melanotos Pectoral Sandpiper
Calidris alpina Dunlin
Calidris ferruginea Curlew Sandpiper
Calidris himantopus Stilt Sandpiper
Tryngites subruficollis Buff-breasted
Sandpiper
Philomachus pugnax Ruff
Scolopacinae
Limnodromus
griseus Short-billed Dowitcher
Limnodromus scolopaceus Long-billed
Dowitcher
Gallinago imperialis Imperial Snipe
Gallinago jamesoni Andean Snipe
Gallinago stricklandii Fuegian Snipe
Gallinago nobilis Noble Snipe
Gallinago undulata Giant Snipe
Gallinago delicata Wilson's Snipe
Gallinago paraguaiae South American Snipe
Gallinago andina Puna Snipe
Tringinae
Phalaropus
tricolor Wilson's Phalarope
Phalaropus lobatus Red-necked Phalarope
Phalaropus fulicarius Red Phalarope
Xenus cinereus Terek Sandpiper
Actitis macularius Spotted Sandpiper
Tringa solitaria Solitary Sandpiper
Tringa incana Wandering Tattler
Tringa melanoleuca Greater Yellowlegs
Tringa semipalmata Willet
Tringa flavipes Lesser Yellowlegs
Tringa glareola Wood Sandpiper
Additional
points:
1. The
above does not deal with the problems of generic limits in the Calidrinae,
e.g., sister relationship of Aphriza
to C. canutus; see Banks (2012). Dick Banks is submitting a proposal to NACC
on this, and I suggest we do not meddle until dust settled.
2. The
sequence from Xenus through T. glareola now conforms to current NACC
sequence.
3. SACC
proposal 548
would split Chubbia from Gallinago, but that would not change the
linear sequence of species.
4. Banks (2012) used Arenariinae Stejneger, 1885, as the
subfamily name, even though Calidrinae Reichenbach, 1849, is older name: “The
family-group name for the turnstones was originally based on the generic name Strepsilas
Illiger, 1811 by Gray (1840), as Strepsilinae (fide Bock 1994:138). Stejneger
(1885:95) introduced the name Arenariinae, based on Arenarius Brisson,
1760, a name with many years priority over Strepsilas, and it thus
became the proper name for the subfamily (see Ridgway 1919:42). For purposes of
priority, Arenariinae dates from 1840 (ICZN 1999, Art. 40.2). The name
Calidridinae was not established until l849 (fide Bock 1994:138) and thus is a
junior synonym of Arenariinae.”
Discussion
& Recommendation: See proposal 552 for my views on the value of using
additional ranks, in this case subfamily, in our classification. I am open to alternative classifications, but
barring viable alternatives, I recommend a YES on this as improving the
information content of our classification.
Literature:
BANKS, R. C.
2012. Classification and nomenclature of the sandpipers (Aves:
Arenariinae). Zootaxa 3513: 86-88.
GIBSON, R.,
AND A. BAKER. 2012. Multiple
gene sequences resolve phylogenetic relationships in the shorebird suborder Scolopaci (Aves: Charadriiformes). Molecular
Phylogenetics and Evolution 64: 66–72.
LIVEZEY, B.
C. 2010.
Phylogenetics of modern shorebirds
(Charadriiformes) based on phenotypic evidence: analysis and
discussion. Zoological Journal of the Linnean Society 160: 567-618.
Van Remsen, October 2012
Comments from Stiles: “A very qualified YES (but note that Chubbia is not reinstated here but will be if 546 passes. Also see
the extensive Raty commentaries on the Scolopacidae in the “TiF Classification”
(whatever this is). Note that here Calidris
is restricted to canutus and a
couple of OW species, and includes Aphriza;
all the smaller Calidris are treated
in Ereunetes. Also, Limicoli is considered to have priority
over Scolopaci (or -inae, for subfamilies).
These points seem worth considering.”
Comments from Pacheco: “YES. Considerando todos os pontos, incluindo os adicionais.”
Comments
from Cadena: “YES (see comments under proposal 552).”
Comments
from Pérez-Emán: “NO. I
like this proposal better than previous ones on the same topic (552 and 553) because
it incorporates some behavioral and morphological issues into the discussion
(in the case of Phalaropes and Turnstones). However, integrating such
information provides an interesting example about how different interpretations
of the same data might result in a different subfamily classification
(discussions in Gibson & Baker (2012) and in this proposal). Moreover, as
pointed out by Remsen, depending on the importance or interpretation given to
branch lengths or node depths in the tree, one can come out with different
subfamily arrangements. Thus, such borderline situations discussed by Remsen
could be more the rule than the exception and might result in non-meaningful
classifications at the subfamily level.”
Comments
from Nores: “YES. This seems
like the best way to go given available evidence.”
Comments
from Jaramillo:
“NO. I realize the subjectivity of this
enterprise, and that Terek Sandpiper is making things difficult here. But the
Gibson Baker paper is but one set of data, and one could find a lot of
morphological, behavioral and other details that clarify that Terek (Xenus) is a Tringine. I think this is
clearly a spurious result here in the genetic data and I do not think we should
really consider this likely, unless some other new and compelling information
came up to put Xenus in with the
phalaropes. My vote would be to recognize the Phalaropes as a subfamily, and
make the decision to leave Xenus out
of that and put it in the Tringines. But subfamily distinctions should of
course tell us about major subdivisions in a family, but these subdivisions
should be informative, to add some structure and logic to the organization
within the family. It is much more structured and logical as well as
informative to have the Phalaropes under their own subfamily!
“Similarly, I
would give the dowitchers their own subfamily. Snipe and woodcock are a good
group; the division between them is relatively deep, but not as deep as between
them and the dowitchers. The dowitchers are very different from snipe and
woodcocks, and maintaining them as separate adds to a more informative and
structured understanding of this family. Other than long bills which ally them
to the snipe/woodcocks, in terms of coloration, vocalizations, migration and
behavior they converge with some Arenariines (sandpipers) and Limosinines
(godwits). They are distinctly different from woodcocks and snipe.
“I know, where
do you draw the line then? Well, I would keep turnstones in with the rest of
the sandpipers. I draw the line there. They are distinctive in many ways, but
fit well within that relatively diverse group. …. although I could be swayed on
this point.”
Comments
from Zimmer: YES. This
seems to be the most sensible arrangement to me, given the current molecular
data. As for the Phalarope question (separate subfamily or not?), I
heartily concur with Van’s statement that P.
tricolor is as much tringine as phalarope with regards to foraging behavior
and foot morphology.”