Proposal (600) to South American Classification Committee
Split Xiphorhynchus ocellatus into three
species, and treat X. pardalotus as
basal to them
Effect
on SACC:
Xiphorhynchus ocellatus would be
split into X. chunchotambo, X.
beauperthuysii, and X. ocellatus.
Xiphorhynchus pardalotus would be
placed immediately before X. chunchotambo.
Background: Genetic data (Aleixo 2002)
indicate that Xiphorhynchus pardalotus, sometimes considered a member of the X. spixii superspecies, and X.
ocellatus are sister species,
as proposed by Zimmer (1934a); Aleixo (2002) found that pardalotus is embedded within taxa currently treated
under X. ocellatus (see Note 21), and this
should be reflected in linear sequence. Derryberry
et al. (2011) found that X. pardalotus was the sister to their single
population sample of X. ocellatus.
New
Information:
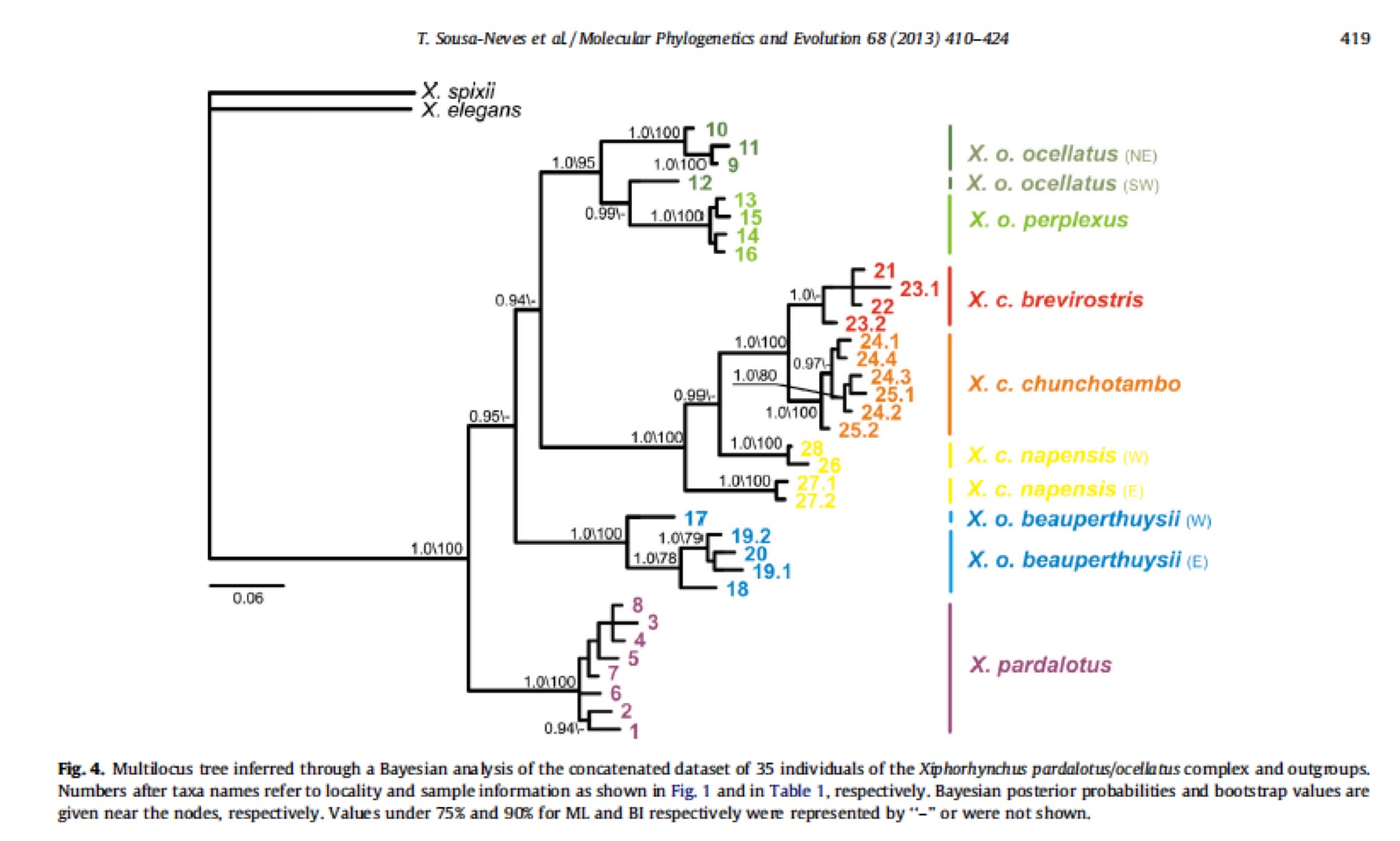
A recently published multilocus phylogenetic analysis on the Xiphorhynchus pardalotus/ocellatus
complex (Sousa-Neves et al., 2013)
recovered a basal position for X.
pardalotus with respect to X.
ocellatus, and the division of X.
ocellatus into three well-supported major clades (PP ≥ 0.95), based on gene
trees and coalescent-based species-tree analyses. The authors sampled 35
individuals of most subspecies recognized to the complex, except X. ocellatus lineatocapilla and X. pardalotus caurensis, whose morphological
diagnoses and validity remain controversial (Marantz et al., 2003). The recovered major clades within X. ocellatus correspond to the proposed
species below:
Xiphorhynchus chunchotambo:
distributed in the lower eastern Andean slopes and adjacent lowland Amazonia
from southern Colombia, eastern Ecuador, eastern Peru, northern Bolivia, and
the southeastern part of the Brazilian state of Acre;
Xiphorhynchus beauperthuysii: distributed north of the Amazon from the western banks of
the Negro River in the Brazilian state of Amazonas through southern Venezuela
(Amazonas), southern Colombia (Amazonas), eastern Ecuador, and northern Peru
east of the Napo river, with an apparently isolated population inhabiting
sandy-soils forests near Jeberos;
Xiphorhynchus ocellatus:
Distributed south of the Amazon in northeastern Peru (east of the lower Ucayali
River) to the Madeira – Tapajós interfluve in the Brazilian states of Amazonas,
Acre, Rondônia, and Pará and also Pando in northern Bolivia.
Analysis/Recommendation: Despite
incongruence between the species and the gene trees – in the mitochondrial gene
tree, X. ocellatus is paraphyletic
with respect to X. pardalotus, for
example (see also Aleixo 2002 and Derryberry et al. 2011) - the coalescent multi-locus and concatenated trees
recovered with overall high statistical support the reciprocal monophyly among X. pardalotus, X. chunchotambo, X. beauperthuysii,
and X. ocellatus, which is consistent
with a high degree of evolutionary independence and reproductive isolation
among these clades (see also Guilherme & Aleixo 2008 discussion on a
contact zone between X. ocellatus and X. chunchotambo in eastern Acre). Even
though a formal quantitative analysis is not presented, vocal types among these
clades are also very distinct (see Xeno-Canto recordings and Sousa-Neves et al. 2013). In fact, significant vocal
variation appears to exist also within at least one of these clades (X. chunchotambo), indicating that
probably additional cryptic species will be uncovered by future studies
(Sousa-Neves et al. 2013).
We therefore strongly recommend a YES
vote on this proposal, which we divide into three parts:
A.
Recognize Xiphorhynchus
chunchotambo as a separate species from X. ocellatus.
B. Recognize
Xiphorhynchus
beauperthuysii as a separate species from X. ocellatus.
C. Move X. pardalotus in the linear sequence to
precede these taxa.
Literature Cited
ALEIXO, A. 2002. Molecular systematics and the role
of the "várzea"-"terra firme" ecotone in the
diversification of Xiphorhynchus woodcreepers (Aves:
Dendrocolaptidae). Auk 119: 621-640.
DERRYBERRY, E., S. CLARAMUNT, G. DERRYBERRY, R. T.
CHESSER, J. CRACRAFT, A. ALEIXO, J. PÉREZ-ÉMAN, J. V. REMSEN, JR., AND R. T.
BRUMFIELD. 2011. Lineage diversification and morphological
evolution in a large-scale continental radiation: the Neotropical ovenbirds and
woodcreepers (Aves: Furnariidae).
Evolution 65: 2973-2986.
GUILHERME, E., AND ALEIXO, A., 2008. Primeiros registros de Xiphorhynchus chunchotambo (Tschudi,
1844) (Dendrocolaptidae) no Brasil. Rev.
Bras. Orn. 16: 44–46.
MARANTZ, C. A., A. ALEIXO, L. R. BEVIER, AND M. A.
PATTEN. 2003. Family Dendrocolaptidae
(woodcreepers). Pp. 358-447 in "Handbook of the Birds of
the World, Vol. 8. Broadbills to tapaculos." (J. del Hoyo et al.,
eds.). Lynx Edicions, Barcelona.
SOUSA-NEVES, T,
ALEIXO, A., SEQUEIRA, F. 2013. Cryptic patterns of diversification of a
widespread Amazonian Woodcreeper species complex (Aves: Dendrocolaptidae)
inferred from multilocus phylogenetic analysis: implications for historical
biogeography and taxonomy. Molecular Phylogenetics and Evolution 68: 410-424.
ZIMMER, J. T.
1934a. Studies of Peruvian birds,
No. 15. Notes on the genus Xiphorhynchus. American Museum Novitates 756: 1-20.
Sidnei Dantas and Alexandre Aleixo
November 2013
Note
from Remsen:
A separate proposal on English names will be needed if A and B pass.
_____________________________________________________________________________________________
Comments from Stiles: “YES on A, B and
C. My only gripe is having to live with
an orthographic horror like beauperthuysii
in Leticia – but I´d already noted the vocal and morphological differences
between this taxon and the birds (chunchotambo)
at the base of the Andes in Caquetá!”
Comments from Pacheco: “[YES on A, B and
C.] O arranjo proposto é fortemente
apoiado nos resultados de um conjunto de estudos.”
Comments from Remsen: “In general,
everyone should take note that this study reveals the importance of multilocus
studies in separating gene trees from species trees.”
A.
“NO. Although I am fairly certain from qualitative descriptions of
vocalizations that chunchotambo
deserves species rank, until these differences are explicitly presented and
quantified, I object to elevating it to species rank. It would take someone about a day of work to
put together a little paper on this taxon’s voice and compare it to ocellatus (s.s.). I think we should adhere to minimum standards
of rigor before making changes to the classification.”
B.
“NO. Without vocal data, all we have no
evidence that beauperthuysii is
anything other than a valid subspecies.”
C.
“YES. Sousa-Neves et al. confirm
conclusively that pardalotus is
misplaced in the linear sequence.”
Comments
from Cadena: “NO. The
multilocus genetic data are highly suggestive of the existence of more than one
species, but they may represent variation existing among allopatric populations
of a single species. I listened to recordings and, sure, they sound different,
but no formal analysis has been published. I thus agree with Van in that we are
not quite there in terms of evidence to propose the three-way split.”
Comments from Zimmer:
“A. NO. Based on vocal differences, I feel pretty
certain that a split is called for, but as noted by Van and Daniel, there has
been no published vocal analysis to demonstrate this. Setting aside the vocal differences, I’m not
enthusiastic about splitting on the genetic data alone, which, although
certainly suggestive, could still reflect variation between long isolated
populations of the same species. I would
welcome a published vocal analysis, which, I anticipate will provide us with
all the justification needed for making the split.
“B.
NO, for similar reasons as in Part A.
“C. YES. Evidence for this seems pretty solid.”