Proposal (639) to South American Classification Committee
Split
extralimital R. l. crepitans group from
Rallus longirostris
Synopsis: The following is the full proposal submitted to NACC on
revising species limits in the Rallus
longirostris/R. crepitans group,
sections of which are not directly relevant to SACC. The key issue is that the
taxon we call Clapper Rail, R.
longirostris, is not the sister species to the North American taxa
currently including in that species. This proposal has been approved and adopted
by NACC. The only taxa present in the
SACC area are members of the Rallus
longirostris group sensu stricto. A
YES vote means that the only change to the SACC list would be the change in
English name of more narrowly defined Rallus
longirostris to “Mangrove Rail”, and a NO vote means no change.
Description
of the Problem:
In
the most recent checklist (AOU 1998:131), there is a discussion of
hybridization between Rallus longirostris
and R. elegans in the eastern and
southern United States. There is a suggestion to merge the entire complex into
a superspecies, but phylogenetic and detailed hybridization studies of the
group have not been published until now (Maley 2012;
Maley and
Brumfield 2013). Taxonomy has always
been difficult in this group given plumage variation, morphologically distinct
allopatric populations, and uncertainty in the degree of hybridization between
populations currently in contact. For example, R. elegans of the eastern US are bright rufous ventrally and breed
in freshwater marshes, whereas R.
longirostris of the eastern US are duller ventrally and breed in saltmarshes.
Very similar allopatric birds of the southwestern US and northwestern Mexico
are bright rufous ventrally and breed primarily in saltmarshes, making their
classification into either species difficult (Olson 1997).
New
Information:
A
phylogenetic study using mitochondrial and nuclear markers found discordance
between genetic relationships and current classification (Maley and
Brumfield 2013). Rallus elegans, as currently recognized, is paraphyletic with
respect to R. longirostris. Genetic
lineages correspond roughly to geography instead of current species limits. The
R. l. obsoletus subspecies group found in California, Arizona, and
northwestern Mexico was discovered to be sister to R. e. tenuirostris of the highlands of Mexico instead of previously
suggested sister relationships to either R.
l. crepitans or R. e. elegans of
eastern North America (Hellmayr and
Conover 1942; Ripley 1977; Olson 1997). Additionally, the
lineages of the R. l. crepitans group
and R. e. elegans, which are known to
hybridize in eastern North America (Olson 1997), are in the same clade (Maley and
Brumfield 2013). This pattern of
hybridization apparently also occurs on Cuba (Olson 1997) between members of these same two lineages (Maley and
Brumfield 2013). This clade also
includes birds from throughout the Caribbean (Fig. 1). Detailed investigations
of hybridization using morphological, ecological, and genetic (mitochondrial
and nuclear) characters in Louisiana reveal that strong selection against
hybrids is likely preventing the fusion of these lineages (Maley 2012). Members
of the nominate R. l. longirostris
group of South America were found to be genetically distinct and sister to
Caribbean and eastern North American birds (Fig. 1B). In the study the authors
were unable to obtain samples of R. l.
longirostris, instead sampling two members of the group R. l. cypereti and R. l. phelpsi. The
following recommendations would remove R.
longirostris from the checklist, because members of this subspecies group
have not been documented in North America.
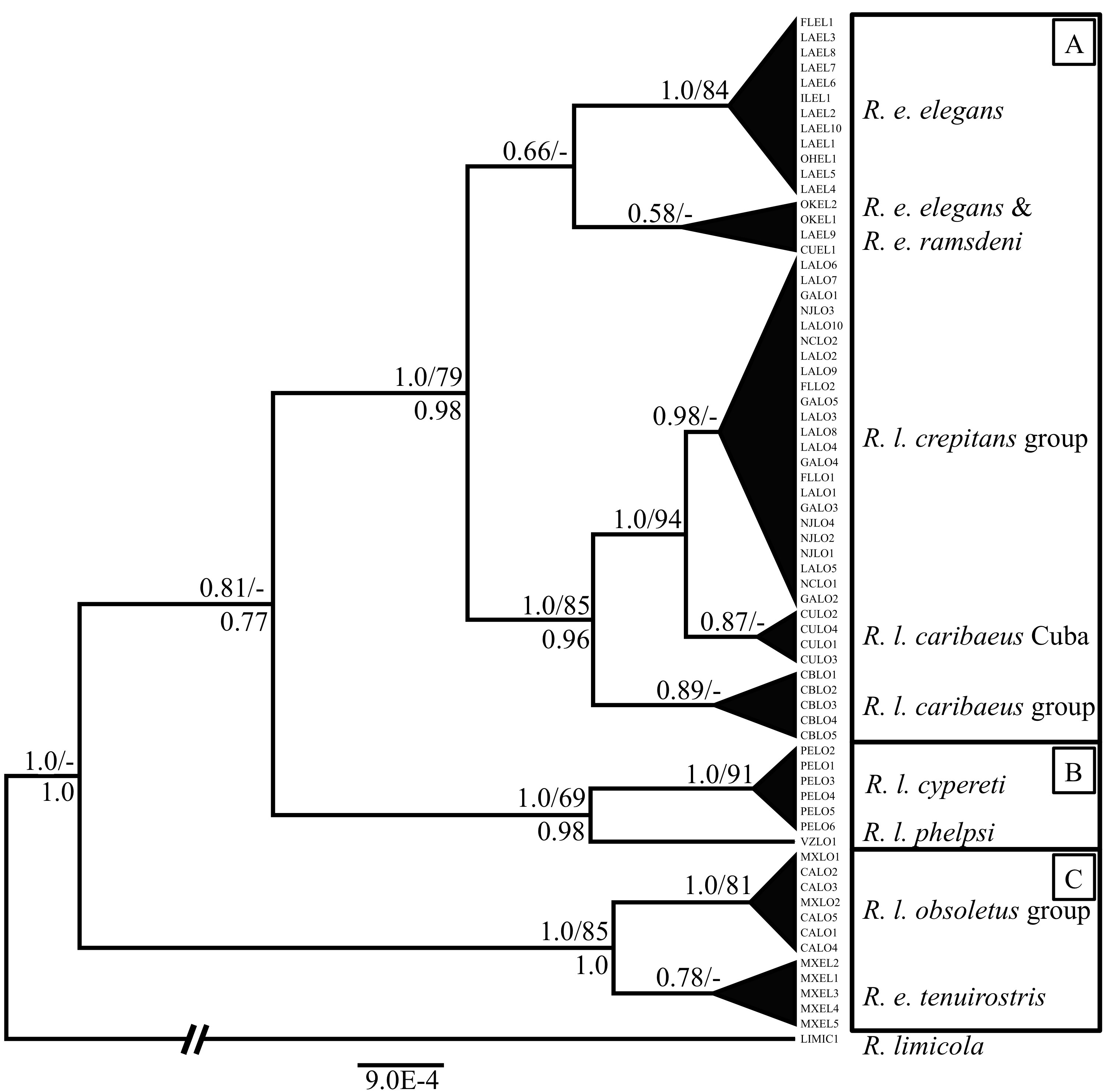
Figure
1. Maximum Clade Credibility gene tree of ND2 inferred in Beast (Drummond and
Rambaut 2007). The labels above
nodes are the posterior probability followed by the bootstrap support value (if
greater than 65) for that node. The labels below nodes are the posterior
probability for that node in the estimate of the species tree; this label is
not included if the value was below 0.95. Each of the three major clades is
outlined and labeled by geography, with clade A comprising eastern North
American and Caribbean birds, clade B comprising South American birds, and
clade C comprising birds of western North America, including Mexico.
Two
members of the complex are in extensive secondary contact in eastern North
America and Cuba, but have not fused despite hybridization (Olson 1997). The morphological and genetic characterization of the
hybrid zone in Louisiana (Maley 2012) found that it is very narrow (~ 4.2 km wide), with
selection against hybrids acting to prevent fusion. These data suggest there is
strong, albeit incomplete, reproductive isolation between these species in
Louisiana. There is no evidence of population genetic structure within R. longirostris in the eastern US, and
very little within R. elegans, so we
extrapolate these results for the entire distribution in the eastern and
southeastern US. Extending these results to the remaining taxa and considering
the differential level of morphological, ecological and genetic divergence
between previously identified subspecies groups, we conclude that at least five
species should be recognized in this complex. This treatment would be
consistent with recent genetic analyses of other members of the family showing
similar levels of divergence (Tavares et
al. 2010; Goodman et al. 2011). The most divergent
clade within the complex, according to mtDNA data, represents a pair of
subspecies groups from both currently recognized species (R. l. obsoletus group and R.
e. tenuirostris). This pair shares the same pattern observed in the birds
of eastern North America, where individuals of one group are relatively smaller
than those of the other and are found primarily in saltmarshes (R. l. obsoletus group), whereas the
other is relatively larger, brighter, and found in freshwater habitats (R. e. tenuirostris,
Olson 1997).
Recommendation:
We
propose species rank for five members of the complex described below. These
taxonomic recommendations are based primarily on two factors: 1) there is
strong but incomplete reproductive isolation between parapatric populations
based on hybrid zone analyses, 2) that each of the species represents a
morphologically and genetically distinct group within the complex that is at
least as distinct from other members of the complex as members that are
currently in contact but showing evidence of reproductive isolation.
We
propose recognizing as a species the nominate form R. longirostris Boddaert, 1783, plus the subspecies phelpsi Wetmore, 1941, margaritae Zimmer and Phelps, 1944, pelodramus Oberholser, 1937, cypereti Taczanowski, 1877, and crassirostris Lawrence, 1871. These
birds are relatively very small, dull-breasted, robust-billed, and restricted
to mangroves (Eddleman and
Conway 1998), which is why we
propose to give them the English name Mangrove Rail. This lineage is
morphologically, genetically, and vocally distinct from all other members of
the complex, and far more distinct from the rest of the complex than the
members that are currently in contact are from one another.
The
second species we propose is R. tenuirostris
Ridgway, 1874, which includes the
population of birds inhabiting the highland freshwater marshes of Mexico.
Individuals are large, very bright rufous ventrally, and have diffuse flank
banding (Meanley 1992). They are found almost entirely within the former Aztec
Empire and are not the only member of the complex found in Mexico; thus we
propose the English name Aztec Rail. They are distinct morphologically,
genetically, and ecologically from their closest relative, in that they breed
exclusively in freshwater marshes as opposed to saltmarshes, which is the same
reproductive isolating mechanism as found in other lineages within the complex.
The
third species we propose is R. obsoletus Ridgway,
1874, which includes the populations that occur along the Pacific Coast of
North America. This species would include the subspecies levipes Bangs, 1899, beldingi
Ridgway, 1882, yumanensis Dickey,
1923, rhizophorae Dickey, 1930, and nayaritensis McLellan, 1927. This group
is characterized by their relatively small body size (although larger than
South American birds), by a bright rufous breast, and by their occurrence
primarily in saltmarshes (Eddleman and
Conway 1998). Because Robert
Ridgway contributed a significant amount of work on the complex, including
describing R. l. obsoletus and R. l. beldingi, we propose the English
name Ridgway’s Rail in his honor. We propose species rank using a comparative
approach: because this lineage is as distinct morphologically, genetically, and
ecologically from its closest relative (R.
e. tenuirostris) as are other members of the complex in contact known to be
reproductively isolated.
The
fourth species in the complex we propose is R.
elegans Audubon, 1834, comprised of two subspecies, R. e. elegans and R. e.
ramsdeni Riley, 1913, while excluding R.
e. tenuirostris (as described above). We propose retention of King Rail as
the English common name. This species is distinct from its closest relatives
ecologically, morphologically, and genetically. Despite hybridization, they are
reproductively isolated from their closest relative in contact, members of the R. l. crepitans group, apparently due to
ecological differences (Maley 2012).
The
fifth species proposed is R. crepitans
Gmelin, 1789, comprised of the eastern North America group of R. l.
crepitans, including the subspecies waynei
Brewster, 1899, scotti Sennett, 1888,
insularum Brooks, 1920, and saturatus Ridgway, 1880, as well as the
birds of the Caribbean and Yucatan, including R. l. caribaeus Ridgway, 1880, pallidus Nelson, 1905, grossi Paynter, 1950, belizensis Oberholser, 1937, leucophaeus Todd, 1913, and coryi Maynard, 1887. These birds are
intermediate in size, and the breast spans a range of colors from very dull,
silvery-gray, to dull rufous. They breed in saltmarshes and salt-meadows of the
Atlantic and Gulf Coasts of North America, as well as mangroves in the Yucatan,
extreme southern Florida, Louisiana, and Texas, and throughout the Caribbean (Eddleman and
Conway 1998). We propose to retain
Clapper Rail as the English common name to avoid confusion. They are distinct
morphologically, genetically, and ecologically. Despite hybridization, they are
reproductively isolated from the other members of the complex they are in
contact with, R. e. elegans and R. e. ramsdeni (Maley 2012).
Literature
Cited:
James M. Maley
and Robb T. Brumfield, August 2013
======================================================
Comments
from Remsen: “YES I reviewed the paper at an early stage
and consider the authors’ taxonomic arrangement to be the one that matches best
the existing data. The elegans and crepitans groups have extensive, multiple contact zones, yet
hybridization is limited by apparent selection against hybrids; thus, they have
to be treated as separate species.
Ripley’s treatment (in his Rallidae monograph) of them as conspecific is
incorrect. Given that the other two
groups, including our R. longirostris
group, are successively more distantly related to the two for which we have a
test of sympatry, the logical taxonomic treatment is to consider them each also
as separate species.
“The Maley-Brumfield name Mangrove
Rail is a good one for reasons stated in their paper. Although the usual policy is to christen each
daughter species from a split with new names to avoid confusion,
phylogenetically this is not a case of splitting a species into two daughters:
in the case of R. crepitans and R. longirostris, they are not close to
being sister taxa, so Maley & Brumfield, followed by NACC, retained the
long-established “Clapper Rail” for the R.
crepitans group. What NACC calls extralimital
R. crepitans is actually irrelevant to SACC, but I mention this here
in case the issue is raised.”
Comments from Zimmer: “YES” to change the
English name of the more narrowly defined Rallus
longirostris to “Mangrove Rail”.
Given the data showing strong selection against hybrids from the various
contact zones in North America, the splits already adopted by the North
American committee appear justified. Because
our South American longirostris-group
is morphologically, genetically, vocally and ecologically distinct from all
other members in the complex, and these distinctions are greater than those
between sympatric/parapatric members of the complex, treatment of the South
American populations as a distinct species seems pretty straightforward. The proposed English name of “Mangrove Rail”
is perfect for these birds.”
Comments from Stiles: “YES. The only part of the proposal that
affects SACC is the English name Mangrove Rail, which is eminently reasonable.”
Comments
from Stotz: “YES. This split is clear. The North American committee spent way too
much time thinking about English names on this one and came up with Mangrove
Rail for the more limited longirostris. I think Mangrove Rail is a good name and so
favor the split and the English name.”