Proposal
(677) to South American Classification Committee
Recognize Systellura longirostris ruficervix,
Systellura longirostris roraimae, and Systellura longirostris decussata as
species
Effect on SACC: Elevate 3 subspecies within Systellura longirostris to species rank.
Background: Band-winged Nightjar Systellura
longirostris is a very widespread, polytypic species, distributed from
northern and Andean Venezuela south to southern Chile and Argentina, and in
eastern Brazil. Currently no fewer than nine taxa are recognized (Cleere 2010,
Dickinson and Remsen 2013). These taxa generally have a similar plumage
pattern, but differ in plumage saturation and size. A few subspecies, such as ruficervix (Cory 1918, Chapman 1923,
1926) and decussata (Cory 1918)
previously were recognized, based solely on plumage variation, as separate species.
Davis (1978) considered ruficervix to
be a species, but it is unclear how many taxa he vocally examined. Cleere
(1998) included all taxa within longirostris,
but at the time the vocalizations of only a few taxa were known. Later Cleere
(2010) recognized both roraimae and decussata as species, "based on
distinct vocalizations and range"; he included ruficervix in longirostris,
but also commented that "review of subspecies needed" across longirostris.
There is
no comprehensive quantitative survey of vocalizations in longirostris; for that matter, the songs of some taxa may not be
known (e.g. mochaensis, which is restricted to a small region in Chile). However, a nice summary of vocalizations of six taxa in this complex,
with sonograms and audio recordings, was prepared by Andrew Spencer; the
included taxa are longirostris, atripunctata, bifasciata, ruficervix, roraimae, and decussata": see xeno-canto.
More recently, the
situation was summarized by Crestol
(2015):
"Across much of its range, the song of Band-winged
Nightjar is a series of thin, high-pitched whistles. This song
variously is described as "a high thin seeeeerp or seeEEEeert
(emphasis varies), squeezed out, rising then down slurred in pitch" (Hilty
2003; subspecies ruficervix); as "a very high-pitched psee-yeet
or psee-ee-eeyt"
(Ridgely and Greenfield 2001b; subspecies ruficervix); as "a
series of high, thin slurred whistles: teeeEEEEuu" (Lane, in
Schulenberg et al. 2010; subspecies ruficervix and atripunctata);
as "a shrill but weak two-syllabled whistle, tse sweeet"
(Jaramillo 2003; subspecies bifasciata); and as "a
single, weak, very high-pitched plaintive whistle, repeated constantly: tseeooeet
... tseeooeet" (Belton 1984; subspecies longirostris).”
http://www.xeno-canto.org/47214
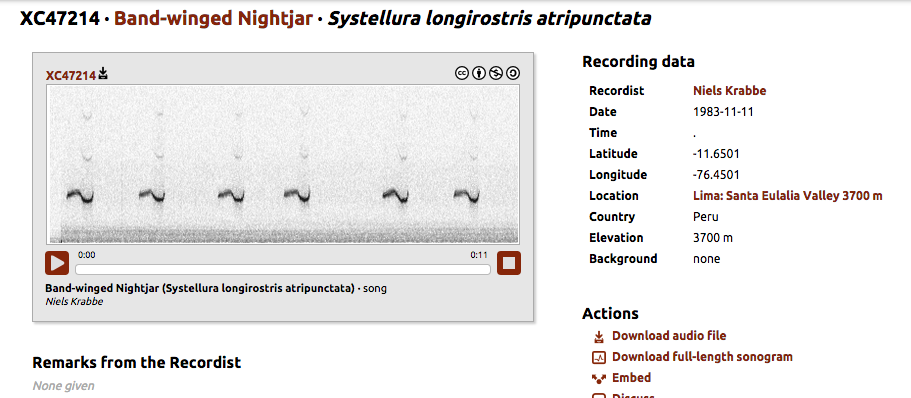
"The song of subspecies roraimae of
the tepuis is very different, and is a series of short, unmodulated, slightly
rising whistles.”
http://www.xeno-canto.org/22131
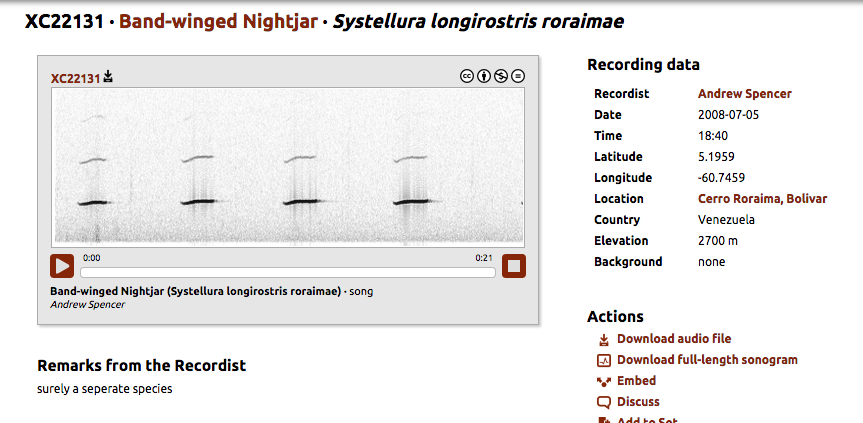
“The song of decussata, of the coastal
lowlands of western Peru and northern Chile, also is distinct: "a loud
series of cueeo notes, reminiscent of [the songs of Common]
Pauraque [Nyctidromus albicollis]
and Scrub
Nightjar [Nyctidromus anthonyi], but
more monosyllabic" (Lane, in Schulenberg et al. 2010)."
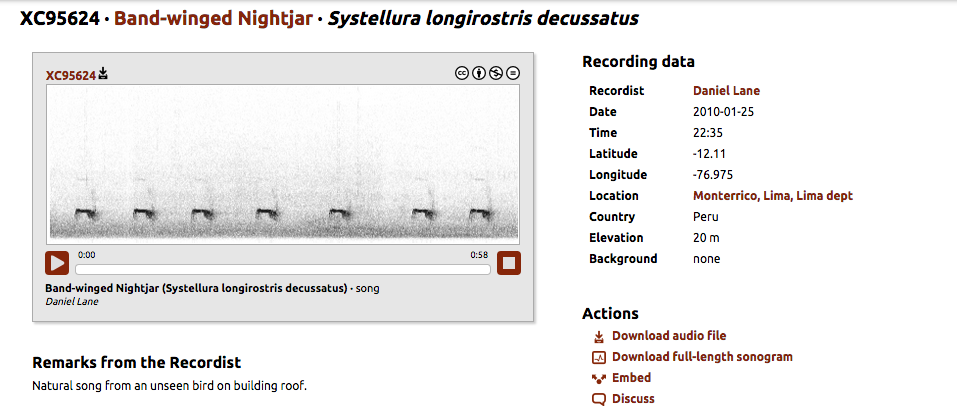
http://www.xeno-canto.org/95624
New Information: The current SACC taxonomy and nomenclature for
Caprimulgidae is based on Han et al. (2010), a phylogenetic analysis of DNA
sequence data (from cytochrome b, and nuclear c-myc and growth hormone genes).
Han et al. included a single example of polytypic longirostris (the subspecific identification was not provided in
the paper, but their sample represents ruficervix).
More recently Sigurdsson
and Cracraft (2014) conducted an independent phylogenetic analysis of New World
caprimulgids, again based on DNA sequence data (NADH dehydrogenase subunit 2,
cytochrome b, RAG-1, and nuclear intron 9 from the aconitase gene). Sigurdsson
and Cracraft also presented the first genetic data for multiple Band-winged
Nightjar taxa, with 18 samples that are identified as representing all but two of
its subspecies: mochaensis of Chile and pedrolimai of
northeastern Brazil.
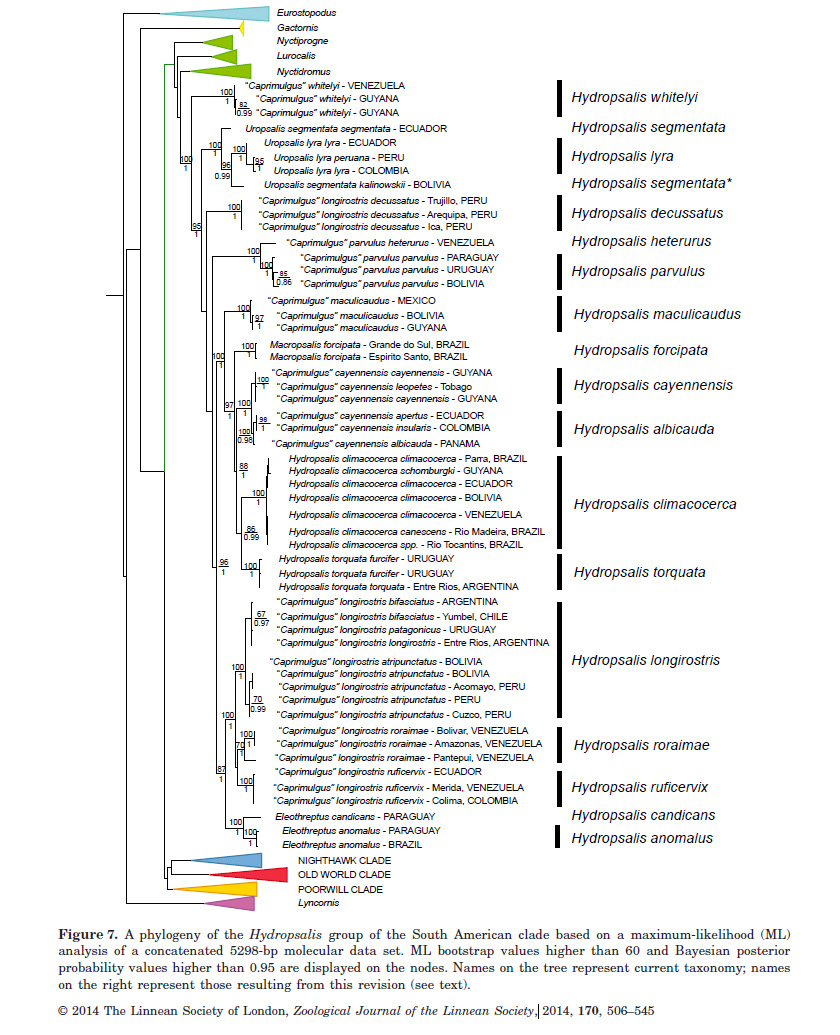
As can be seen from their Figure 7, most taxa
of longirostris form a clade, which in turn has two subunits, one
composed of a group of taxa (atripunctatus, bifasciatus, longirostris,
and patagonicus) representing a wide swath of the geographic range of
the species, and another that pairs two taxa (ruficervix and roraimae)
that occur in montane areas (Andes and tepuis) of northern South America; these
two taxa are darkest in plumage.
A more surprising result
is that the traditional species longirostris is highly polyphyletic, as decussata
is distantly related to other taxa included in longirostris. Instead, it
is basal to the clade that includes the following genera recognized by SACC: Setopagis,
Hydropsalis, Macropsalis, Hydropsalis, Systellura, and Eleothreptus
(!).
Based on the above, the
following actions can be contemplated:
A)
recognize decussata as a species
B)
recognize both ruficervix and roraimae as separate species, based
on the genetic distinctions from the clade atripunctatus, bifasciatus,
longirostris, and patagonicus, and the vocal distinctions between ruficervix
and roraimae
Recommendation: Separation of decussata
as a species is mandatory, as the phylogeny presented by Sigurdsson and
Cracraft makes clear that it is not conspecific with the rest of the longirostris group. Based on the above
data sets, we also recommend a "yes" vote on Part B, for elevating
both ruficervix and roraimae to species level.
If these
pass, we need to think about English names. Cory (1918) and Cleere (2010) used
the name "Tschudi's" for decussata;
we have no objections to this name, and adopting it saves us the trouble of
thinking further about the issue. Cleere (2010) adopted the name "Tepui
Nightjar" for roraimae. which
again is a good name. Davis (1978) made the logical suggestion of "Rufous-naped
Nightjar" for ruficervix.
Obviously ruficervix is not the only
"rufous-naped" nightjar, but then, longirostris is not the only "band-winged" nightjar. So,
we propose that a "yes" vote for Parts A or B also constitutes assent
to our proposed English names, unless otherwise specified in comments with each
vote. If a committee member doesn't like our proposed names, then suggest
something else! If necessary, a separate vote can be held later on alternative
English names.
Finally,
note that decussata clearly cannot be
classified in the genus Systellura.
If SACC votes to recognize decussata
as a separate species, then a second proposal will be submitted on a generic
classification for decussata.
Literature Cited:
Belton, W. 1984. Birds of Rio Grande do Sul, Brazil. Part 1.
Bulletin of the American Museum of Natural History 178: 369-636.
Chapman, F.M. 1923. Descriptions
of proposed new birds from Panama, Venezuela, Ecuador, Peru and Bolivia.
American Museum Novitates number 67.
Chapman, F.M. 1929. Descriptions
of new birds from Mt. Roraima. American Museum Novitates number 341.
Cleere, N. 1998. Nightjars: a guide to
the nightjars, nighthawks, and their relatives. Yale University Press, New
Haven, Connecticut.
Cleere N.
2010. Nightjars, potoos,
frogmouths, oilbirds and owlet-nightjars of the world. Old Basing, UK: WILDGuides Ltd., Parr House.
Cory, C. B. 1918. Catalogue of birds of the
Americas. Part II, number 1. Field Museum of Natural History
Zoological Series volume 13, part 2, number 1.
Crestol, S. 2015. Band-winged
Nightjar (Systellura longirostris),
Neotropical Birds Online (T. S. Schulenberg, editor). Cornell Lab of
Ornithology, Ithaca, New York.
Davis, LI. 1978. Acoustic evidence of relationships in
Caprimulginae. Pan American Studies 1: 22-57.
Dickinson, E.C., and J.V. Remsen, Jr.
(editors). 2013. The Howard and Moore complete checklist
of the birds of the world. Fourth edition. Volume 1. Non-passerines. Aves
Press, Eastbourne, United Kingdom.
Han,
K.-L., M.B. Robbins, and M.J. Braun. 2010. A multi-gene estimate of phylogeny
in the nightjars and nighthawks (Caprimulgidae). Molecular Phylogenetics and
Evolution 55: 443-453.
Hilty, S.L. 2003. Birds of Venezuela.
Second edition. Princeton University Press, Princeton, New Jersey.
Jaramillo, A. 2003. Birds of Chile.
Princeton University Press, Princeton, New Jersey.
Ridgely, R.S., and P.J. Greenfield.
2001b. The birds of Ecuador: field guide. Cornell University Press, Ithaca, New
York.
Schulenberg, T.S., D.F. Stotz, D.F.
Lane, J.P. O’Neill, and T.A. Parker III. 2010. Birds of Peru. Revised and
updated edition. Princeton University Press, Princeton, New Jersey.
Sigurdsson,
S. and J. Cracraft. 2014. Deciphering the
diversity and history of New World nightjars (Aves: Caprimulgidae) using
molecular phylogenetics. Zoological Journal of the Linnean Society. 170:506–545.
Tom Schulenberg and Mark Robbins, July
2015
=========================================================
Comments
from Zimmer: “This
proposal seems to be split into two parts.
Part A) Recognize decussata
as a species: YES. On this, the genetic data are clear. Part B) Recognize both ruficervix and roraimae as separate species:
YES. Genetic differences support
splitting these two from the main clade that includes nominate longirostris, and vocal and range
considerations would support treating ruficervix
and roraimae as separate from one
another. It has long been suspected that
“Band-winged Nightjar” encompassed multiple species. Adopting this proposal would be a good start
toward resolving some of the confusion, and provide a framework for redirecting
focus on the status of some of the remaining taxa under the longirostris-umbrella. The proposed English names seem reasonable.”
Comments from Stiles:
“A)
YES. At the very least, decussatus is
clearly not a member of the clade containing the rest of longirostris.
“B) YES. Vocalizations and genetics support splitting S. roraimae and ruficervix from longirostris,
and this also implies that atripunctatus and
the other southern races will also constitute at least one other species,
assuming that the Argentina sample is not nominate longirostris (although here, I assume that vocal data plus more
thorough genetic sampling are needed
to clarify this group).”
Comments from Areta:
“A) YES. The situation with decussata is
different, as it is not part of the PRESUMED longirostris complex, and
it appears to have been sufficiently well sampled.
“B) “NO. The taxonomy of Systellura
longirostris has long been known to be problematic and inaccurate. However,
I see several problems in adopting the proposed taxonomy in Sigurdsson &
Cracraft (2014).
“1)
Nominate longirostris, atripunctatus, and bifasciatus/patagonicus
have distinctive vocalizations (as diagnostic as those of other taxa afforded
species rank in their proposed taxonomy). Lumping them under a single species
is not recommended if we are to split the others, as it would mean having an
internally inconsistent Systellura.
“2)
Their single sample of nominate longirostris from Entre Rios (Argentina)
is very likely a migratory bifasciatus/patagonicus and not a true longirostris.
I speculate this based on known winter migration of bifasciatus/patagonicus,
and the lack of confirmed records of nominate longirostris in Argentina.
Thus, strictly speaking, we do not know what longirostris is in terms of
phylogenetic placement. This being said, since I fear that nominate longirostris
was not sampled, we do not know if longirostris indeed deserves to be in
the genus Systellura (type species Stenopsis ruficervix).
“3)
Subspecific distinctions of patagonicus and bifasciatus are
contentious, and they might not even be diagnosable (but more work is needed on
this). So it is surprising to find subspecific identification attached to the
bird in Uruguay, which must also be a winter visitor.
“4)
We (Thomas Valqui, Chris Witt and I) have been working for some time on a paper
on this complex, analyzing more recordings of all taxa, and with a better
genetic sampling. At present, even when there is disperse data clearly
indicating that current species limits are wrong, there is not a careful and
thorough characterization of variation and discreteness of vocalizations of the
involved taxa. Thus, I'd rather postpone a decision until such a work is
published.
“5) I
must stress that if we make a decision on this proposal based on xeno-canto
recordings, then a cascade of other proposals without critical proper review
must pass too. We all know of several groups with erroneous taxonomy and could
put 'quick' proposals together compiling disperse data to support a different
taxonomic treatment. I am of the view that solid taxonomic works require
careful scrutiny and attention to detail. I thus do not feel comfortable with
deep taxonomic changes in nightjars when the goal of a paper was not to
evaluate species limits, when the rationale for subspecific identification of
taxa is not made clear, and when relevant natural history information is not
included in the arguments. The work by Sigurdsson & Cracraft (2014) is a
great contribution to our understanding of phylogenetic relationships in
nightjars. But I do not think it contains enough information to solve this
puzzle.
Comments from Remsen:
“A) YES, for reasons outlined by others.
B) NO – I am strongly persuaded by Nacho’s reasoning.”
Comments
from Pacheco: “A) YES,
in accordance with what has been presented.
B) NO. After
the careful arguments provided by Nacho, I prefer to wait for more evidence to
decide about the specific limits within that complex.”
Comments from Nores: “A: YES. B: YES, especially with the
vocal difference between ruficervix
and roraimae, which are together in a
separate clade.”
Comments
from Jaramillo: “A. YES.
The form decussata/decussatus is
clearly not part of this group, based on molecular data. It is also the most
vocally divergent. If this proposal does go through, I would NOT be happy with
Tschudi’s Nightjar. I would rather choose an ecological/habitat name. Is Desert
Nightjar taken in the Old World?.
“B. YES. Although this may be
piecemeal and incomplete based on Nacho’s comments, I think that it would be
best to make a decision on these two (ruficervix
and roraimae) now. We have the data,
and it looks good. I doubt that Nacho’s work will not agree in the separation
of these two forms. I am guessing that their work may consider more splits in
the longirostris/bifasciatus/atripunctatus
etc. groups. But that can happen later, and I don’t see that it impacts the
current decision. I agree with Nacho that the Sigurdsson & Cracraft paper
does not “contain enough information to solve this puzzle” but it does contain
enough to solve some of the puzzle. So I think a YES on part B of this proposal
is defensible. By the way, I am unsure
of where exactly mochaensis is found,
but assume that Chiloe Island is part of the distribution. These birds sound,
to my ears at least, just like birds from Central Chile. They also respond to
recordings of birds from Central Chile.”
Comments
from Stotz: “
A. decussatus YES
B. split ruficervix and roraimae
YES
“I understand
that this is not the final story on this complex, but it seems like recognizing
these is not really affected by the fact that there may still be further splits
to do within longirostris. There are other taxa where we have recognized
splits, while still having complexes that have not been fully worked out within
the unit. An example of that would be
the recognition of Tolmomyias traylori,
despite the fact that are almost certainly multiple species within T. sulphurescens.”
Comments from Cadena: “677A. YES. 677B. NO. These sonograms
do look different and songs sound rather distinct, but are they representative
of variation existing within each of the groups and consistent across
geography? I think we cannot tell until data are properly (i.e. quantitatively)
analyzed with careful geographic sampling and such analyses are published.
Also, although this does not matter much given the proposal, the proposal
states that the longirostris clade
has two “subunits” with one corresponding to the montane taxa ruficervix and roraimae, but I note there is no support for these two taxa forming
a clade.”