Proposal
(689) to South American Classification Committee
Transfer
Helmeted Woodpecker from Dryocopus to
Celeus
Effect on SACC: This would transfer
the Helmeted Woodpecker Dryocopus galeatus
to Celeus.
New
Information:
Since Peters (1948), the Helmeted Woodpecker has been placed in the genus Dryocopus because of its similarity in
plumage to members of that genus, in particular, the Lineated Woodpecker D. lineatus. Short (1982) noted plumage and structural
characters of galeatus that were
shared with Celeus, but he continued
to maintain it in Dryocopus. The development of a robust molecular-based
phylogeny and comparative analysis of vocalizations for Celeus (Benz and Robbins 2011) coupled with field experience by
several observers suggested that the Helmeted Woodpecker was not a Dryocopus. This precipitated obtaining genetic material
of galeatus that clearly place this unique woodpecker in Celeus (Benz et al. 2015; Figure 5):
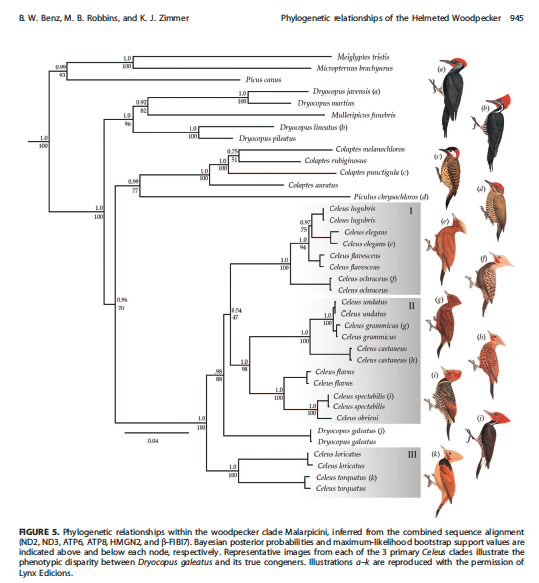
Recommendation: The molecular data
clearly establish that the Helmeted Woodpecker is a member of Celeus. The strikingly similar plumage
pattern shared with sympatric Lineated Dryocopus
lineatus and Robust Campephilus robustus woodpeckers is
likely due to mimicry, although that hypothesis needs to be tested (Benz et al.
2015).
Literature cited:
Benz, B.W. and M.B. Robbins. 2011. Molecular Phylogenetics, Vocalizations, and Species Limits
in Celeus Woodpeckers (Aves:
Picidae). Molecular Phylogenetics and Evolution 61:29–44.
Benz, B.W., M.B. Robbins, and K.J.
Zimmer. 2015. Phylogenetic relationships of the Helmeted Woodpecker (Dryocopus galeatus): A case of
interspecific mimicry? Auk 132:939-950.
Peters 1948. Check-list of birds of the
world. Vol. 6. Museum of Comparative Zoology, Harvard University Press,
Cambridge, Massachusetts.
Short, L.L. 1982. Woodpeckers of the
World. Delaware Museum of Natural History, Greenville,
Delaware.
Mark
Robbins, Kevin Zimmer, and Brett Benz, November 2015
=========================================================
Comments from Remsen: “YES. Fascinating
result, strong data, easy decision.”
Comments by Areta: “YES. A long awaited (and suspected)
phylogenetic result, which is almost exactly replicated by Lammertink et al.
(2015). Benz et al. (2015) and Lammertink et al. (2015) differ in the exact
placement of galeatus although all other species sampled agree in their
phylogenetic relationships. Where will we place galeatus? Lammertink et
al. (2015) placed it as sister to flavus-obrieni-spectabilis,
while Benz et al. (2015) placed it as sister to all sampled Celeus
excepting torquatus-loricatus. I incline towards Benz et al.
(2015) proposal given their better taxon and gene sampling.”
Comments
from Stiles: “YES – a
neat study, fascinating result (but consistent with the mimicry hypothesis for Campephilus-Dryocopus parallel changes
in coloration!)”
Additional
comments from Brett Benz:
“To address Aretha’s concerns regarding the phylogenetic position of galeatus
within Celeus, I downloaded the ND2 sequences generated by Lammertink
et al (2015) from GenBank to compare it to those of Benz et al (2015) and Dufort
2016. The Benz et al ND2 sequences
differed from that produced by the Dufort study by only 2 base pairs, whereas
the sequences generated by Lammertink differed by 6 and 12 base pairs
(~0.5 to 1.1%), with the later sequence containing 5 unique amino acids
at positions that are otherwise invariable within all of Celeus.
Although visual inspection of the Lammertink ND2 sequences revealed several
instances where base pairs were likely miscalled resulting in sequentially
shifted codon positions (a pattern indicative of a dirty trace file from the
sequencer), this is unlikely the source of differing topologies between
studies, as the Dufort super matrix approach also recovered a different
topology for Celeus and placement of galeatus. Both the Lammertink and Dufort studies
partitioned sequence data by gene, whereas Benz (2015) partitioned
the mtDNA data by codon position, allowing for greater model specificity at
conservative 2nd positions. Thus, marker selection and data-partitioning
strategies likely account for the topological differences among these
studies. The artificially high node scores (all 100%) of the Lammertink et
al. study illustrate the importance of careful model selection and attention
to methodological detail.”
Comments
from Stotz: “YES. This is a stunning finding, but the
supporting data are strong.”
Comments
from Zimmer: “As a
co-author on the Benz et al 2015 paper, this one gets an expected YES vote from
me. The molecular data clearly establish
phylogenetic relationships that vocal, morphometric, behavioral and ecological
characters had all suggested, all of which were obscured by the remarkable
convergence of plumage patterns. I’ll
never forget spending several minutes looking at my first galeatus at Iguaçu Falls NP back in the early 1990s and remarking
to my tour group that despite the plumage pattern, this bird struck me as being
a Celeus, a conviction that was only
strengthened the first time that I heard one vocalize. Not surprisingly, Ted Parker (ICBP Red Data
Book, 1992) had already noted some morphological, vocal and behavioral
similarities to Celeus, as, apparently,
had Short before him. Nice to get this
fascinating case recognized.”
Comments from
Pacheco: “YES. Surprising,
but very well supported by the data.”
Comments
from Jaramillo: “YES –
Not really surprising because people in the field have felt this was the case
for a long time. So it is great to see a different dataset confirm this notion,
which seemed preposterous given the plumage.”