Proposal (720)
to South American
Classification Committee
Treat White-breasted Wood-Wren Henicorhina leucosticta as two or
more species
Background
Information
Henicorhina leucosticta inhabits lowland humid forest from eastern Mexico to
central Colombia and the Chocó region of northwestern Ecuador, and also in the
western and northern Amazon Basin (Fig. 1). H. leucosticta varies subtly
in plumage throughout its distribution, and is classified into a dozen or so
subspecies based largely on plumage differences (Table 1). H. leucosticta also
shows geographic variation in vocal traits including pitch and repetition of
song style (Brewer 2001).
Two phylogenetic studies have examined
geographic patterns of mtDNA in H. leucosticta (Dingle et al 2006,
Lelevier 2008). Both studies identified three major clades—one in Central
America, a second in the Chocó of northwest Ecuador, and a third in the Amazon
basin. These three clades are deeply divergent, with estimated 7–12% mtDNA
sequence divergence between clades (Lelevier 2008). Additional sampling in
Panama in the Lelevier study revealed that birds in the Darien of eastern
Panama are sister to the Amazon clade. These Darien birds could be considered
part of the Amazon clade or a distinct “Darién” clade; for clarity when
discussing different populations, we follow the latter course in this proposal.
The geographic distribution of the Darién clade remains uncertain, though it
contacts i) the Central American clade in central or eastern Panama and ii) the
Chocó clade, likely somewhere in western Colombia. More recently, Smith et al
(2014) included H. leucosticta in their large phylogeographic analysis
of Neotropical lowland taxa, and, using genomic data, recovered very similar
relationships to those previously identified using mtDNA (see screenshot
below).
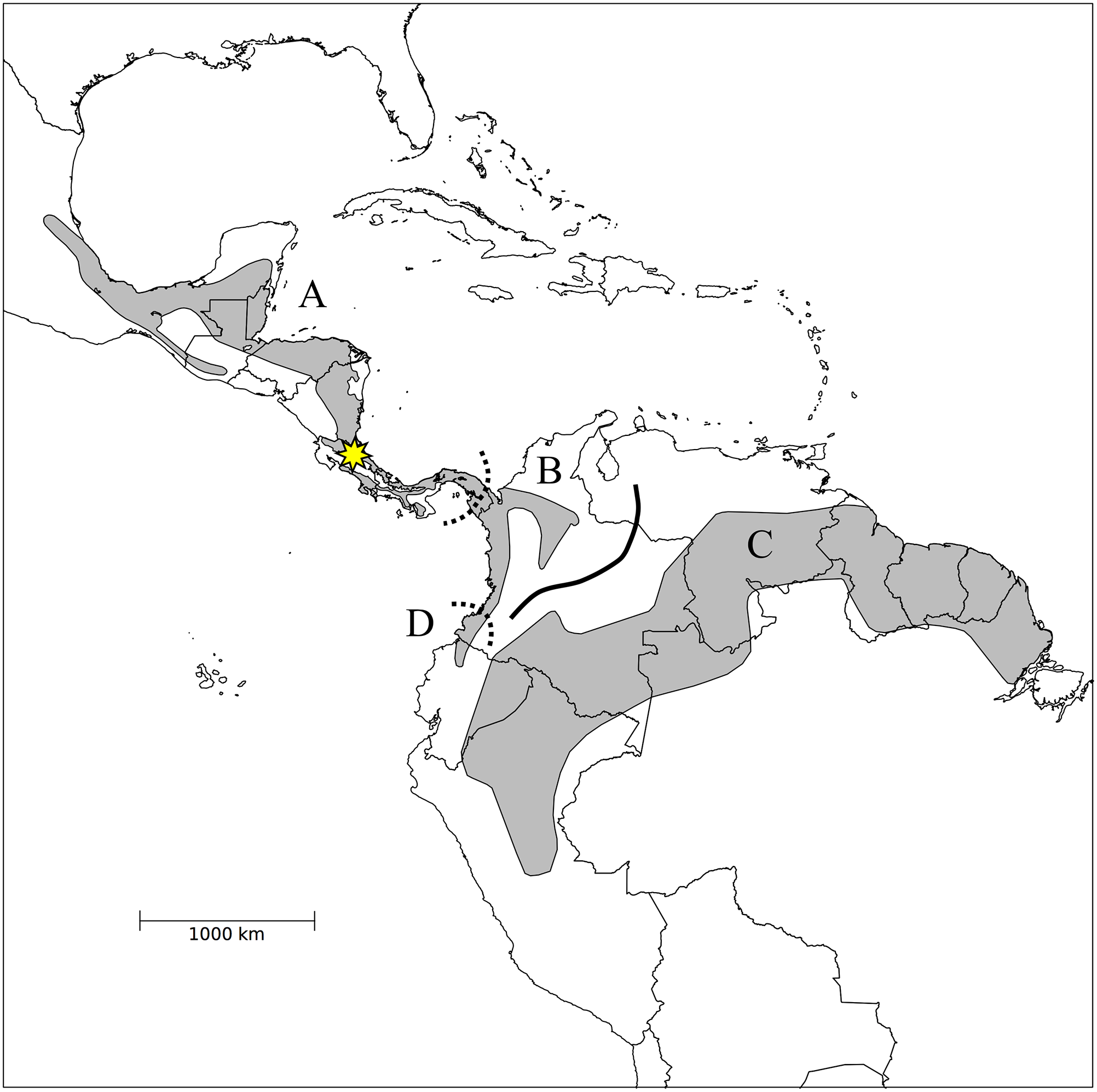
Fig. 1 from Pegan et al 2015. Range
map of H. leucosticta illustrating the distributions of four
mitochondrial clades (A = Central American, B = Darién, C = Amazon, D = Chocó;
the precise geographic distribution of the Darién clade is unknown and marked
in dashed lines). The location of La Selva Biological Station in the Caribbean
lowlands of Costa Rica, where fieldwork for the Pegan et al. study was
completed, is marked by a star. Map from BirdLife International (2014).
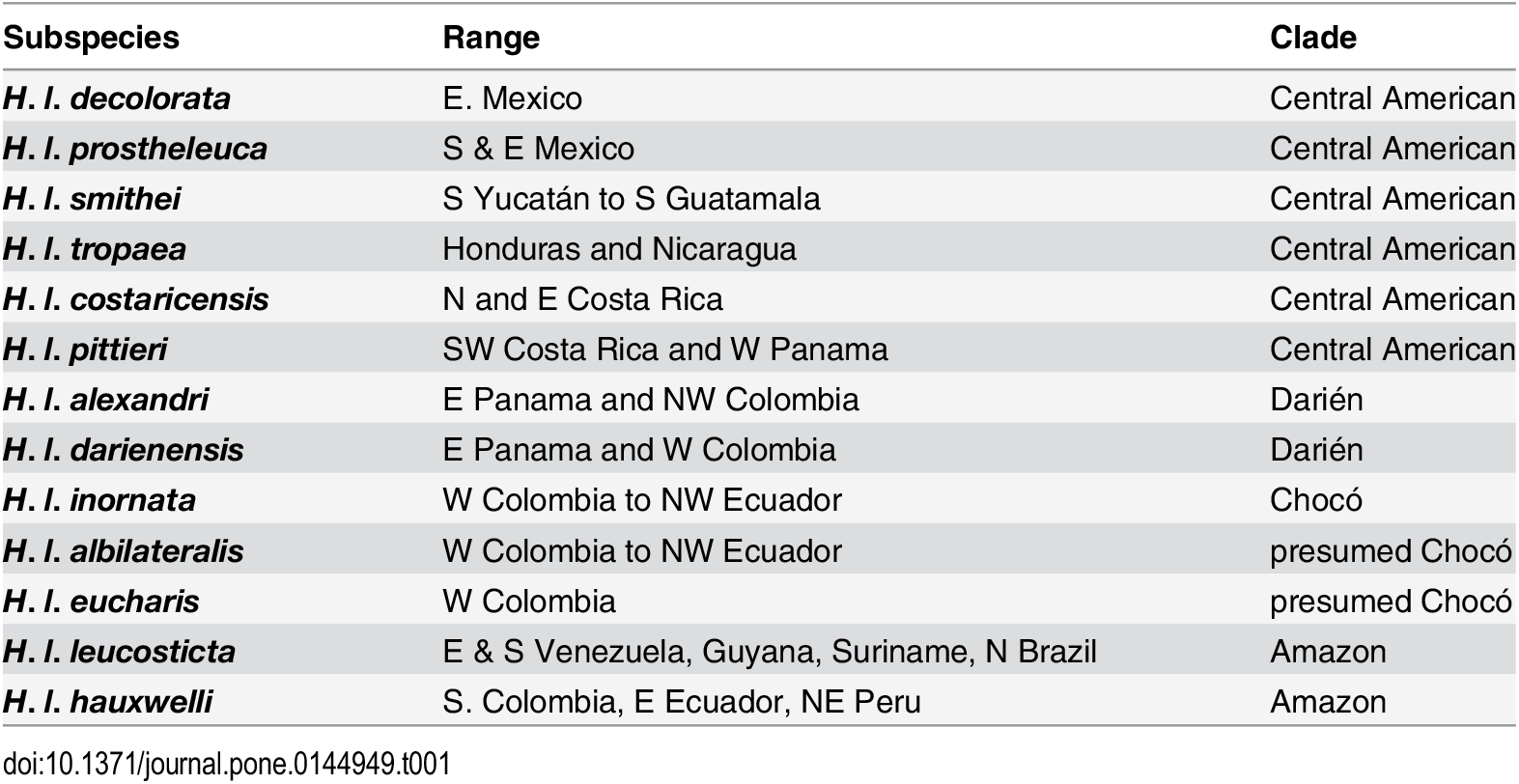
Table 1 from Pegan et al. 2015.
Summary of subspecies and their clades based on Lelevier (2008). Birds from western Colombia are presumed to be within the
Chocó clade pending further evidence.
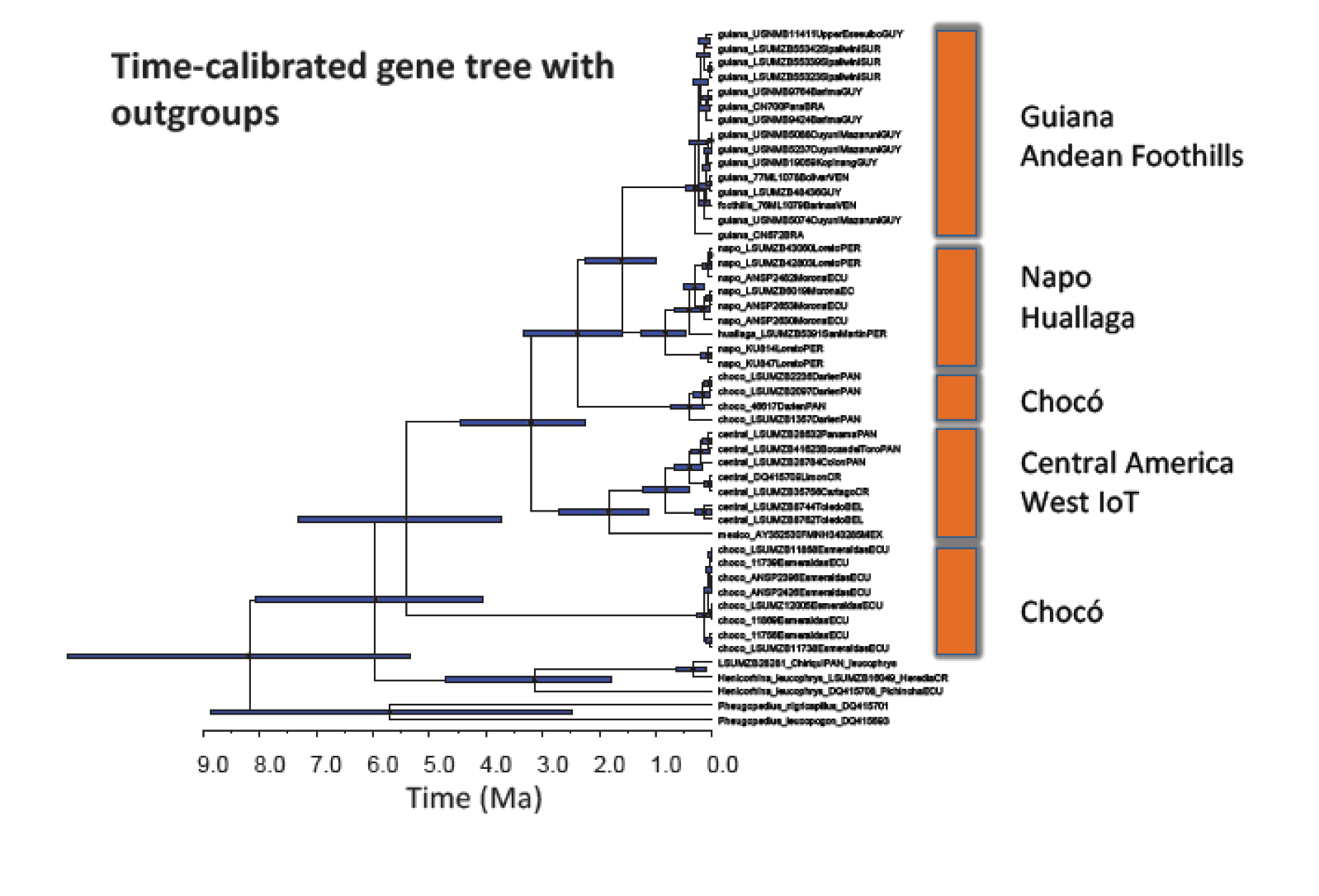
Close
up of Figure S10 from Smith et al. 2014 showing time calibrated gene tree. Note
that Smith et al. designate the clade sister to the
Amazonian populations found in the Darien region of eastern Panama– that we are
calling “Darién” in this proposal— as “Choco”.
New
information:
Recently, Pegan et al (2015) used
vocal trait analysis and field playback experiments to assess premating
reproductive isolation in song in H. leucosticta. While the utility of
these approaches are well understood for suboscines, divergence in
vocalizations between allopatric populations can also be relevant to species
limits in oscines.
Vocal trait analyses: Pegan et al.
analyzed eight song variables (song length, total note count, mean number of
notes per syllable, mean note rate, peak frequency, mean note high frequency,
mean note low frequency, and total song frequency range) for a representative
song from a total of 49 recordings from three H. leucosticta clades (n =
19 from the Costa Rican population, n = 10 from the Chocó population in
northwestern Ecuador and western Colombia, and n = 20 from the Amazon
population), then visualized population differences in vocal traits using the
first two axes from a principal components analysis (PCA) that quantified
variation across the nine vocal characters (see Fig. 2).
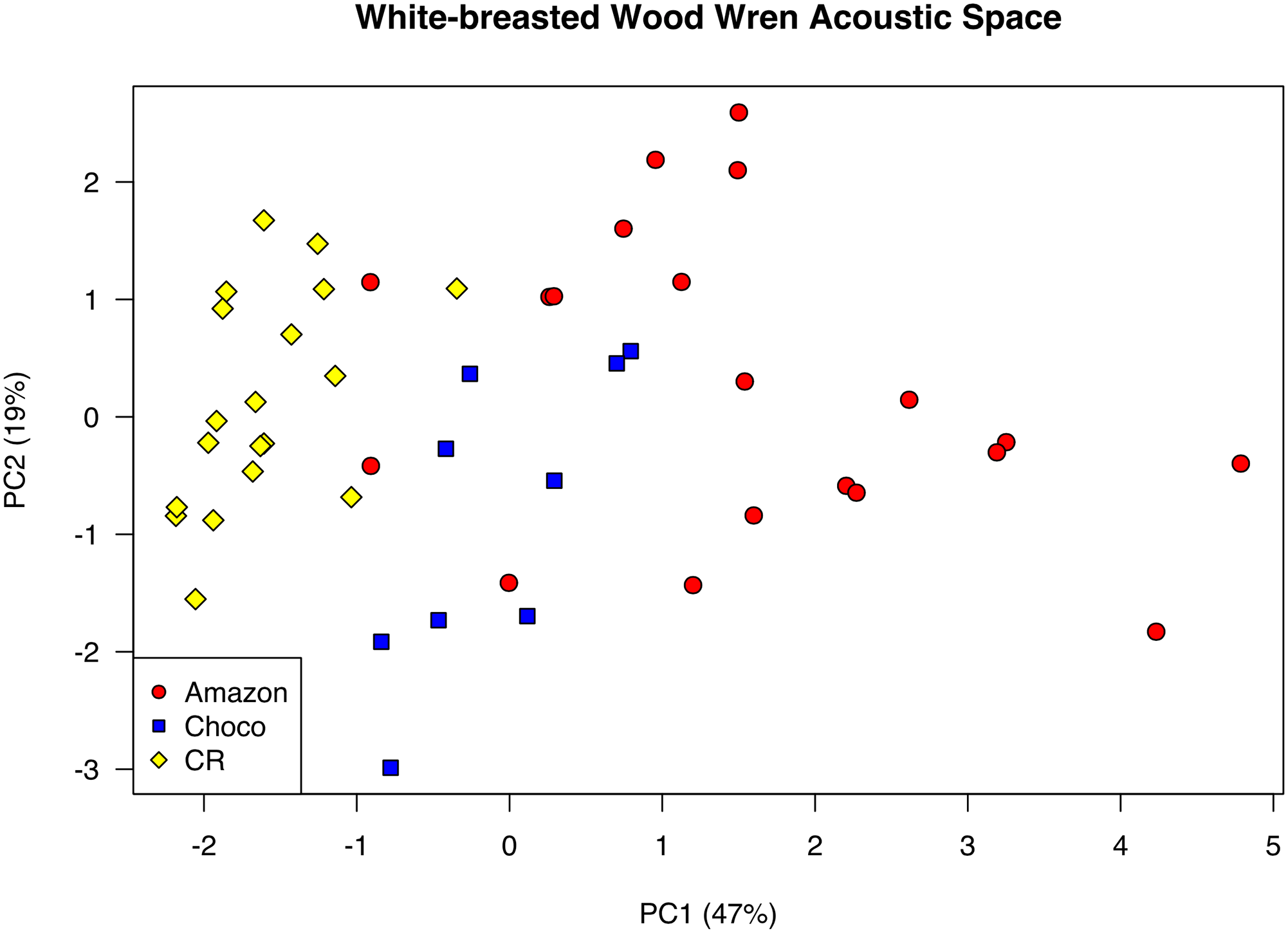
Fig. 2 from Pegan et al. 2015. PCA
plot of acoustic space.
Larger values on PC1 represent songs
that are longer, that have more notes, and that cover a wider range of
frequencies. Larger values on PC2 represent songs with faster note rates and
smaller frequency ranges. Although variation in these vocal characters is
continuous across the entire sample, individual populations are differentiated
in acoustic space. In the figure below yellow represents birds from Costa Rica;
Amazonian populations are shown in red; and blue denotes
samples from Chocó (northwestern Ecuador).
doi:10.1371/journal.pone.0144949.g003
Field playback experiments: Pegan et
al. (2015) conducted field experiments on a single population of H.
leucosticta at La Selva Biological Station, Heredia province, in the
Caribbean lowlands of northeastern Costa Rica. Each experiment contained two
treatments; a sympatric treatment of local Costa Rican song playback (H. l. costaricensis) and an allopatric treatment of song playback from a
different population. One experiment (hereafter the “Amazon experiment”) used
playback of vocalizations from WBWWs from Amazonia as the allopatric treatment
(H. l. hauxwelli) whereas the other (hereafter the “Chocó experiment”) used
playback of Chocó clade vocalizations (H. l. inornata) as the allopatric treatment. Trials for these two experiments
were carried out on different territories at La Selva and analyzed
independently.
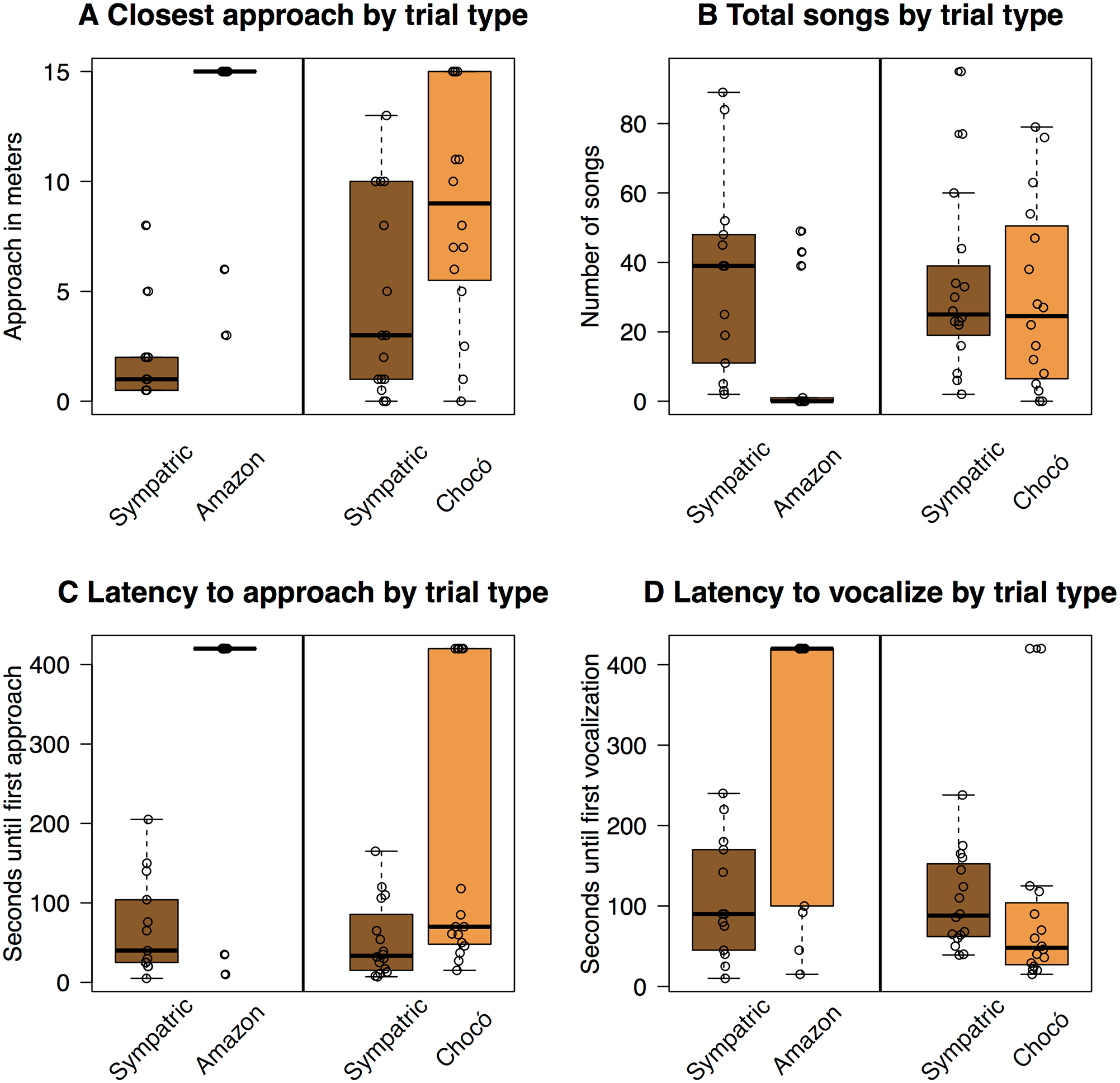
Fig. 3 from Pegan et al 2015.
Behavioral response to playback experiments.
Responses to the Amazon experiment are
on the left and responses to the Chocó experiment are on the right. Latency to
approach (A), closest approach to speaker (B), total number of songs (C) and
latency to vocalize (D). Boxplots illustrate median (horizontal black bar),
first and third quartiles (boxes), and minimum and maximum values (points and
dotted lines). Raw data are plotted as points in front of the boxplots, with
points offset slightly to better display values. Sympatric treatments elicited
aggressive responses (fast approaches, close approaches, many songs, low
latency to vocalize) in both experiments. Allopatric treatments elicited
asymmetric responses: individuals typically responded aggressively to playback
of Chocó songs but not to playback of Amazonian songs.
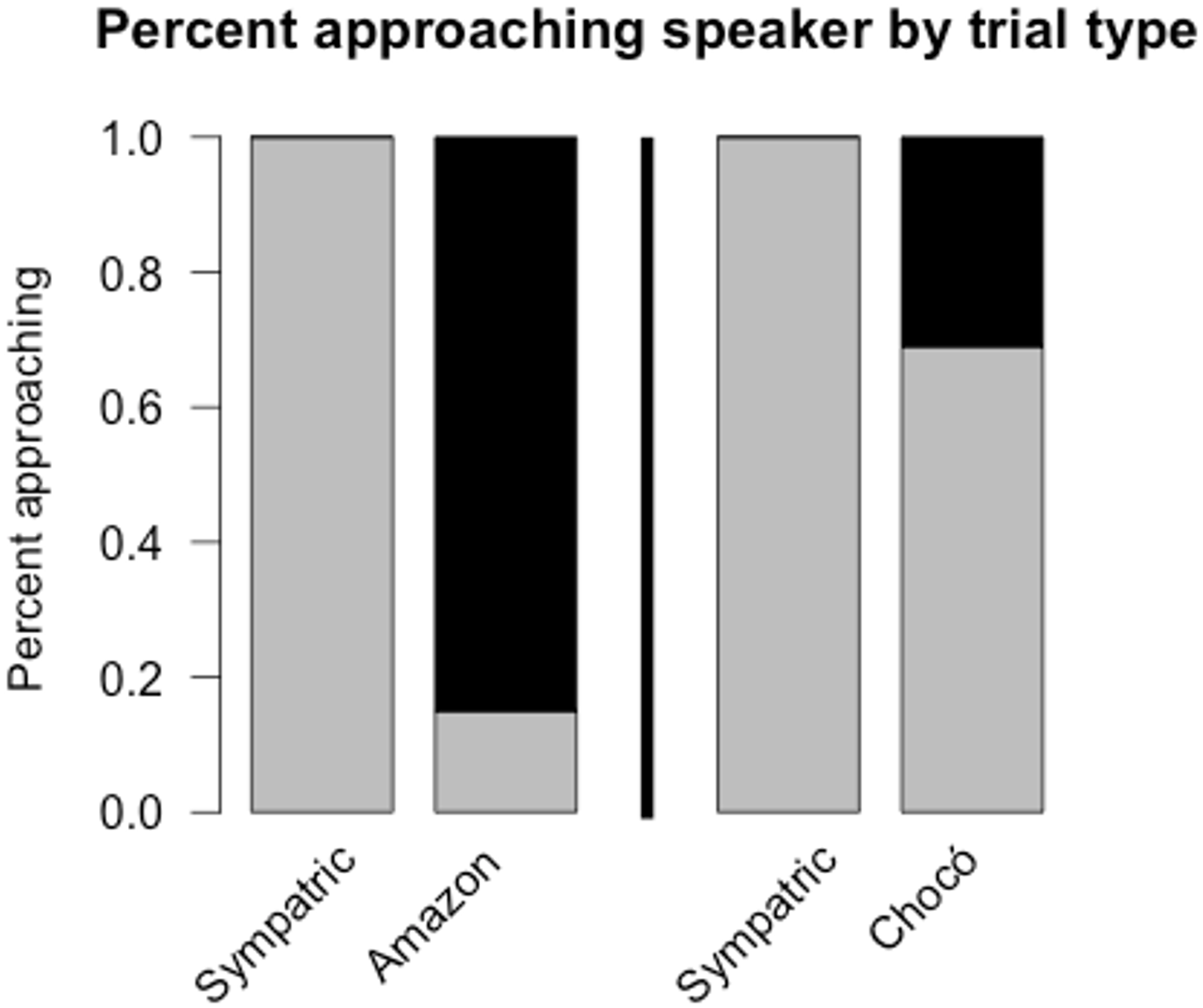
Fig. 4 from Pegan et al. 2015. Percent
of individuals approaching the speaker in each experiment.
Responses to the Amazon experiment are
on the left and responses to the Chocó experiment are on the right. Note that
sympatric trials serve as positive controls (wrens approached the speaker in
all sympatric trials).
Additional
points to consider:
Pegan et al. did not consider the
Darien clade in analysis of vocal traits or playback experiments, as the
geographic limits of this clade beyond the Darien region in eastern Panama
remain unclear and there are few recordings from the Darien in Panama. Though
not quantified, birds from the Darien are vocally similar to Amazonian
population, black-capped (as are Amazonian birds), and genetically sister to
the combined Amazonian populations (e.g. see Smith et al. topology). In
addition, recent (March-April 2016) fieldwork indicates that H. leucosticta in Costa Rica largely
ignore songs from the Darien clade.
Pegan et al. did not conduct
reciprocal playback analyses. Responses to playback are not always symmetrical
between populations (Colbeck et al 2010)—Pegan et al. interpret the lack of
Central American response to Amazonian playback as evidence of premating
reproductive isolation between these two populations, but did not have the
opportunity to measure how Amazonian birds respond to vocalizations from
Central American birds. Given the marked differences in vocal traits, however,
it seems reasonable to infer that Amazonian birds are unlikely to respond
strongly to Central American vocalizations.
Third, Pegan et al. tested a
territorial response rather than a mate choice response. In general, research
suggests that females are more discriminating than males when it comes to
songs, implying that if males discriminate between songs, as we demonstrate, it
is likely that females do as well (Cure et al 2010, Danner et al 2011, Nelson
et al 2004, Searcy 1990, Searcy and Brenowitz 1988). All vocal trait analyses
and playback experiments aimed at determining species limits make the
assumption that territorial songs and mate choice are correlated, and though
this is generally accepted as a valid assumption, it may not be true in all
cases, especially in species with multiple song types where certain types may
be used for territoriality and others for mate attraction.
The playback experiments presented in
Pegan et al (2015) show that H. leucosticta in
Central America do not appear to recognize H. leucosticta song
from the Amazon clade as conspecific (but, interestingly, do generally
recognize song from the even more genetically divergent Choco clade as
conspecific). These field experiments echo the results of a vocal trait
analysis that indicates songs from the Amazonian and Central American clades
are mostly (but not entirely) separated in acoustic space. Genetic data also
shows deep divergence between the Central American and Amazon clades. In sum,
available evidence suggests that Amazon and Central American populations of H. leucosticta represent distinct biological species and
should not be classified as conspecific.
Recommendation:
The suggestion that multiple
biological species lurk within H. leucosticta is
hardly novel, and we expect future work (e.g., in the zones where the Darien
clade contacts the Central American and Choco clades, also within the Amazonian
population) will lead to further insights relevant to H. leucosticta taxonomy.
We note that H. leucosticta as currently defined is paraphyletic with
respect to H. leucoptera, and that this complex remains paraphyletic in
Options 1, 2 and 5 (but not 3 and 4).
We support Option 2 (or Option 1,
though we believe Option 2 is preferable) as a first step forward in improving
the species-level taxonomy of this complex group. Option 3 also is a reasonable
interpretation of the available evidence. Options 4 and 5 are offered for the
sake of completeness, but in our view are not well-supported at this time.
Nomenclature can be considered at a future date if votes favor one of the
splitting options.
Option 1: Split Amazonian population
of H. leucosticta from all populations found west of the Andes. This
reflects the finding of Pegan et al. that H.
leucosticta in Costa Rica largely ignore song from the Amazonian
population.
Option
2: Split Amazonian and Darien populations
of H. leucosticta from all populations found west of the Andes. This
reflects the finding of Pegan et al. that H.
leucosticta in Costa Rica largely ignore song from the Amazonian population
and also the close relationship between Amazonian and Darien populations; these
two clades are sister groups (Smith et al 2014), share plumage traits (both
black capped; Choco and Central American birds have brown caps), and are
similar acoustically (unpublished playback experiments also indicate that Costa
Rican H. leucosticta largely ignore song from the Darien population).
Option 3: Split H leucosticta into three – the Central American clade, the Choco
clade, and the Amazonian + Darien clade. There is no current behavioral
evidence indicating premating reproductive isolation between the Choco and
Darien clades, but they are highly genetically divergent and it would not be
surprising if the two clades behave as biological species wherever they come
into contact in western Colombia.
Option 4: Split H leucosticta into four– the Central American clade, the Choco
clade, the Amazonian clade and the Darien clade. This arrangement was proposed
to be the species tree of this complex by Smith et al (2014).
Option
5: No action for now – retain the
status quo.
References
BirdLife International and NatureServe. Bird
species distribution maps of the world. BirdLife International, Cambridge, UK
and NatureServe, Arlington, USA; 2014
Brewer D. Wrens,
Dippers, and Thrashers. New Haven: Yale University Press; 2001.
Cadena CD, Caro LM, Caycedo PC, Cuervo AM, Bowie RCK, & Slabbekoorn H. Henicorhina
anachoreta (Troglodytidae), another endemic bird species for the
Sierra Nevada de Santa
Marta, Colombia. Ornitología Colombiana
2015;15.
Colbeck G, Sillett T, Webster M. Asymmetric
discrimination of geographical variation in song in a migratory passerine.
Anim Behav. 2010;80: 311–318. doi: 10.1016/j.anbehav.2010.05.013
Curé C, Mathevon N, Mundry R, Aubin T.
Acoustic cues used for species recognition can differ between sexes and sibling
species: evidence in shearwaters. Anim Behav. 2010;84: 239–250. doi:
10.1016/j.anbehav.2012.04.039
Danner JE, Danner RM, Bonier F, Martin PR,
Small TW, Moore IT. Female, but not male, tropical sparrows respond more
strongly to the local song dialect: implications for population divergence. Am
Nat. 2011;178: 53–63.
doi: 10.1086/660283. pmid:21670577
Dingle C, Lovette IJ, Canaday C, Smith TB.
Elevational zonation and the phylogenetic relationships of the Henicorhina wood-wrens. Auk. 2006;123: 119–134.
doi: 10.1642/0004-8038(2006)123[0119:ezatpr]2.0.co;2
Lelevier M. Phylogeography of three widespread
Neotropical avian taxa: Rufous-tailed Hummingbird, White-breasted Wood-Wren and
Anthracothorax mangos. Ph.D. Thesis, University of Alaska Fairbanks.
2008. Available: https://scholarworks.alaska.edu/handle/11122/5182
Nelson DA, Soha JA. Male and female
white-crowned sparrows respond differently to geographic variation in song. Behaviour.
2004;141: 53–69.
doi: 10.1163/156853904772746600
Pegan TM, Rumelt RB, Dzielski SA, Ferraro MM,
Flesher LE, et al. Asymmetric Response of Costa Rican White-Breasted Wood-Wrens
(Henicorhina leucosticta) to Vocalizations from Allopatric Populations. PLoS ONE
2015; 10(12) e0144949. doi: 10.1371/journal.pone.0144949
Reif J, Jiran M, Reifová R, Vokurková J,
Dolata PT, Petrusek A, et al. Interspecific territoriality in two songbird
species: potential role of song convergence in male aggressive interactions. Anim
Behav. 2015;104: 131–136. doi: 10.1016/j.anbehav.2015.03.016.
McEntee JP. Reciprocal territorial responses
of parapatric African sunbirds: species-level asymmetry and intraspecific
geographic variation. Behav Ecol. 2014;25: 1380–1394. doi: 10.1093/beheco/aru136.
Searcy WA. Species recognition of song by
female red-winged blackbirds. Animal Behaviour. 1990:40; 1119–1127. doi: 10.1016/s0003-3472(05)80178-2
Searcy WA, Brenowitz EA. Sexual differences in
species recognition of avian song. Nature. 1988;332: 152–154. doi: 10.1038/332152a0
Seddon N, Tobias JA. Character displacement
from the receiver’s perspective: species and mate recognition despite convergent
signals in suboscine birds. Proc Biol Sci. 2010;277: 2475–83. doi: 10.1098/rspb.2010.0210.
pmid:20375056
Smith,
BT, McCormack, JE, Cuervo, AM, Hickerson, MJ, Aleixo, A, et al. The drivers of
tropical speciation. Nature 2014;515(7527):406-409.
Benjamin Freeman and Teresa Pegan, May 2016
_________________________________________________________________
Comments from Stiles: “NO. This proposal
is part of a larger problem for which the solution is not yet at hand. The
authors present evidence that the northern form responds differently to its own
songs vs. those of an Amazonian form (although the playback experiments were
not reciprocal), suggesting that the two could be considered separate
species. However, the combination of
vouchered specimens with song recordings and DNA samples from across the wide
range of the leucosticta complex is
yet too incomplete to fully resolve the puzzle. The form or forms intervening
between the northern and Amazonian birds are especially poorly sampled. My only
personal contribution to the debate is a bird of this group that I recorded in
Puerto Solano, NW Colombia that sounded so different from the birds I knew in
Costa Rica that I didn’t even recognize its song as that of a wren until I
played it back and had the bird screaming it back at me from 3m away! So, I’d
put their suggestion on hold for now: suggesting species status for the birds
at the extremes of the distribution of the leucosticta
complex without knowing what lies between does not seem justified to me at
this time.”
Comments
from Areta: “NO. As underscored by Gary, I think that
there are still too many missing pieces in this puzzle. I would like to see
more thorough reciprocal playback experiments using larger sample sizes and
better acoustic characterizations of populations being tested. This paper is
one step forward in the taxonomy of Henicorhina leucosticta, but more
need to be taken before taking a robust decision. With the data at hand (and
with the playback experiments suggesting stronger responses to more distantly
related populations), we cannot place unsampled (or sampled!) populations
confidently to species-level taxa.”
Comments from Robbins: “NO. I fully
concur with comments made by Gary. Much
more information is needed before we start subdividing this complex.”
Comments
from Pacheco: “NO. The
consensus is that there is most likely more than one species in this complex. But I agree with the views of colleagues that
the evidence is still incomplete for a good decision.”
Comments from Remsen: “YES on Option 2. Evidence from these excellent studies is
overwhelming that that a minimum of two species are represented, and I see no
real reason to delay at least a 2-way spit until other studies further
subdivide those two (other than likely problems with English names as daughters
are further subdivided).”
Comments from Claramunt: “NO. I agree that more than one species
may be involved, but the data published so far are not sufficient to redefine
species limits. The mitochondrial data is suggestive of population structure
but by itself is not sufficient to demonstrate the existence of more than one
lineage. I’m not impressed by the deep mitochondrial “divergences”, either.
Such deep “divergences” could be just old coalescences, which seem common in
widespread lowland tropical species, a consequence of large, old,
geographically structured, and stable populations. Note also that data in Smith
et al. (2014) is also mitochondrial, not genomic/nuclear; therefore, the
congruence among datasets is expected (they are all mitochondrial). Congruence
among mitochondrial and nuclear datasets would have been a strong test for the
existence of more than one lineage, but we don’t have that information so far.
Variation in external/morphological traits is apparent but an analysis is
needed to determine how these traits vary, whether they vary in a continuous or
discrete fashion, whether they define coherent groups, and whether those groups
coincide or conflict with the mitochondrial groups. Thus, much more data and
work is needed to test whether there are one or more lineages in this complex.
Also critical to determine species limits in this group would be to analyze
more samples from the Chocó, where the Darien and Chocó populations could be in
contact.
“The vocal data are fairly
inconclusive. First, variation in song characteristics seems continuous instead
of defining discrete groups. As for their geographic position, songs from the
Chocó occupy an intermediate position in the multivariate space, suggesting
that vocal geographic variation may be clinal. Playback experiments further
highlight the potential for the clinal variation in songs to allow for species
recognition and interbreeding throughout the range of this species: birds from
Costa Rica responded strongly to songs from the Chocó, and to a lesser degree
to Amazonian vocalizations. Therefore, it is possible that neighboring
populations do recognize each other as conspecific. Then, the relevant test for
assessing reproductive isolation (and species status under the “Biological”
species concept) would be to analyze a more dense chain of neighboring
populations to see if there is some sort of break in species recognition. If
there is no break and neighboring populations recognize each other as
conspecific across the entire range, there is no intrinsic barrier to gene
flow. “Isolation tests” performed on widely allopatric populations (e.g. Costa
Rica versus Amazonia) are irrelevant if they are connected by intermediate
populations with intermediate characteristics and gene flow can occur across
the entire range.”
Comments from Zimmer: “NO”. The evidence presented certainly suggests
that more than one biological species is involved, and my own field experience
has likewise indicated as such, at least as regards vocal differences. However, I am troubled by the fact that the
conclusions of the vocal analysis and playback experiments rest on a comparison
of populations near the geographic extremes of the rather extensive
distribution, without a clear picture of what’s happening in between. Problems that I see include the
following: 1) The playback experiments were
one-way, not reciprocal, so that we know the response of the Costa Rican birds
to songs of the Amazonian clade and the Chocó clade, but we know nothing of how
those populations respond or fail to respond to playback of the Costa Rican
birds. 2) The Darién clade is not included in the experiments or the vocal
analysis. 3) The geographic limits of
the Darién clade are unknown. 4) There
are two named subspecies from western Colombia and nw Ecuador whose inclusion
in the Chocó clade is only presumed. 5) In
playback trials, Costa Rican birds did not respond to playback of Amazonian
birds, but did respond strongly to playback of Chocó birds, which are even more
genetically divergent from the Central American clade than are the Amazonian
birds. On top of all of this, I am even
more concerned about the vocal analysis itself.
According to the proposal, Pegan et al (2015) analyzed eight song
variables, which is great. But I can’t
help but wonder how they addressed the issue of song-types. My experience with Henicorhina is that each population of each species has a song
repertoire that includes multiple song-types, and that the size of the song
repertoire may be individually variable.
I vividly remember an experience with a White-breasted Wood-Wren from my
early days of Neotropical birding at Palenque (Chiapas, Mexico). I tape-recorded natural (unsolicited) songs
of a leucosticta, and played those
songs back, in an attempt to see the bird.
In response to playback, the wren in question immediately switched to a
2nd, distinctly different, song-type. I recorded song-type #2 and played that back,
which stimulated the wren to switch to a 3rd song-type. This was repeated until I recorded song-type
#5 and played it back, only to have the wren recycle song-type #1! It would seem that any vocal analysis
comparing only one song-type from Population A to one song-type from Population
B could produce seemingly diagnosable characters that are, in fact, merely
reflecting sampling bias. Conversely,
failure to control for song-type discrimination (i.e. including more than one
song-type in the same analysis) within the sample from a single population,
could introduce all kinds of variation in the measurement of individual
characters, such that populations could appear to overlap in characters, when,
in fact, it’s a case of apples to oranges.
Given that these are oscines that, at least to a certain extent, learn
their songs, I would like to see a bit more rigor applied to the vocal
analysis, and would like to have a lot more information from the central
portions of the geographic distribution (rather than just from the extremes)
before attempting to assess species-limits, even though I believe the authors
are correct in their assertions that at least two different species are
involved.”