Proposal
(723) to South American Classification Committee
Revise the linear sequence of Orders
This proposal would revise the higher-level
linear sequence as follows:
(a) Move Strigiformes to precede Trogoniformes
(b) Move Accipitriformes to precede Strigiformes
(c) Move Eurypygiformes and Phaethontiformes to precede Sphenisciformes
(d) Reverse the linear sequence of Podicipediformes
and Phoenicopteriformes
(e) Move Pterocliformes and Columbiformes to follow Podicipediformes
(f) Move Cuculiformes, Caprimulgiformes, and Apodiformes to follow
Columbiformes
(g) Move Gruiformes and Charadriiformes to precede Eurypygiformes
(h) Move
Opisthocomiformes to precede Gruiformes
Background:
The
higher-level phylogeny of birds has been addressed over the past 25 years by
genetic studies using various types of data and taxon sampling (e.g., Sibley
and Ahlquist 1990, Groth and Barrowclough 1999, Fain and Houde 2004, Hackett et
al 2008, McCormack et al. 2013, Kimball et al. 2013, Jarvis et al. 2014,
Burleigh et al. 2015, Suh et al. 2015, Prum et al. 2015). The trees produced by
these studies have been at odds with traditional ideas of avian phylogeny in a
variety of ways, some of which have been widely adopted. For example, it is now
generally recognized that the Galliformes and Anseriformes are sister taxa and
together (as Galloanseres) are the sister group to all other extant
non-paleognath birds, or Neoaves (Sibley and Ahlquist 1990, Groth and
Barrowclough 1999, et al.). Non-traditional results among the Neoaves have been
slower to gain acceptance, primarily because they had been found in only a
single study (e.g., in Hackett et al. 2008; Fig. 1) or had been weakly
supported. However, recent phylogenomic studies, especially Jarvis et al.
(2014; Fig. 2) and Prum et al. (2015; Figs. 3 and 4), have provided strong
support for several non-traditional results obtained in one or more previous
studies. Several clades in the higher-level phylogeny of Neoaves are now
reasonably well supported, including large core waterbird and core landbird
clades. The core waterbird clade consists of the Gaviiformes, Sphenisciformes,
Procellariiformes, Ciconiiformes, Suliformes, and Pelecaniformes, and the core
landbird clade consists of the Accipitriformes, Strigiformes, Coliiformes,
Leptosomiformes, Trogoniformes, Upupiformes, Bucerotiformes, Coraciiformes,
Piciformes, Cariamiformes, Falconiformes, Psittaciformes, and Passeriformes.
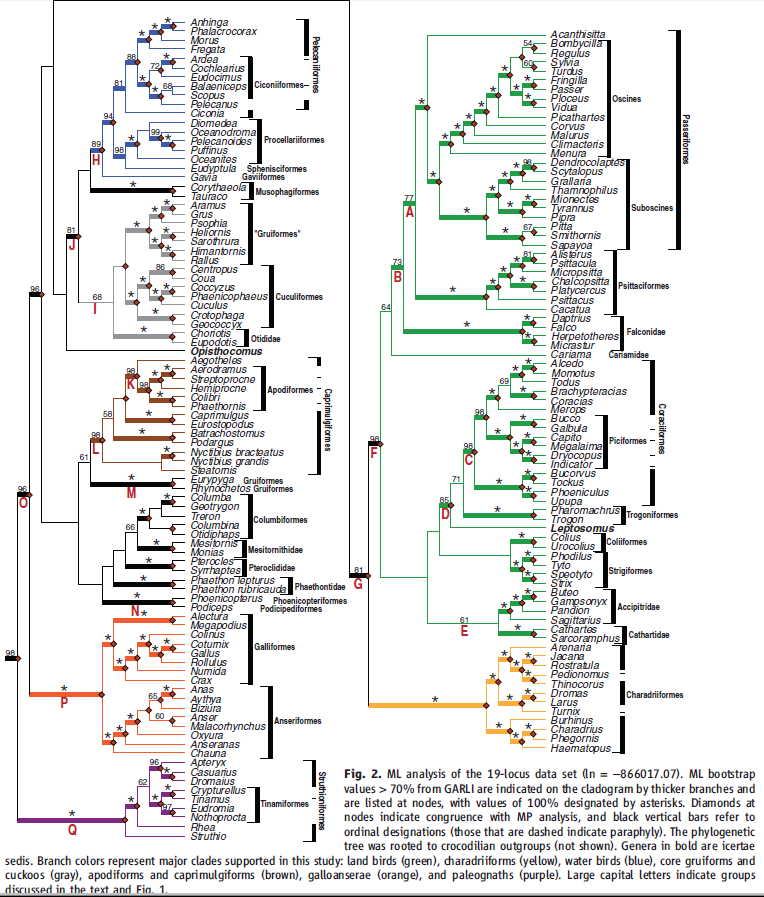
Figure
1. Maximum-likelihood phylogeny from Hackett et al. (2008).
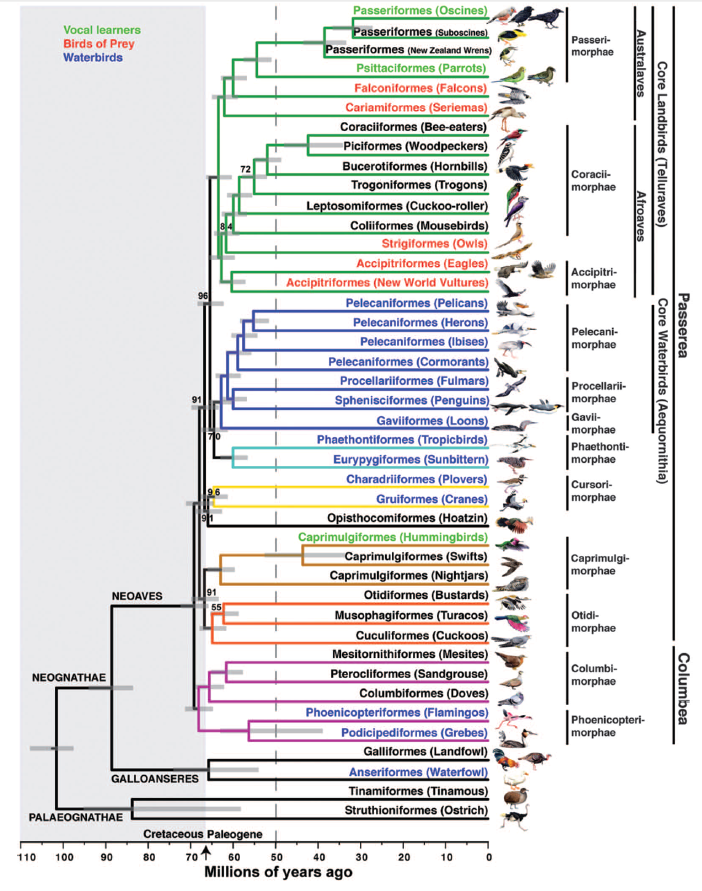
Figure
2. Maximum-likelihood phylogeny from Jarvis et al. (2014).
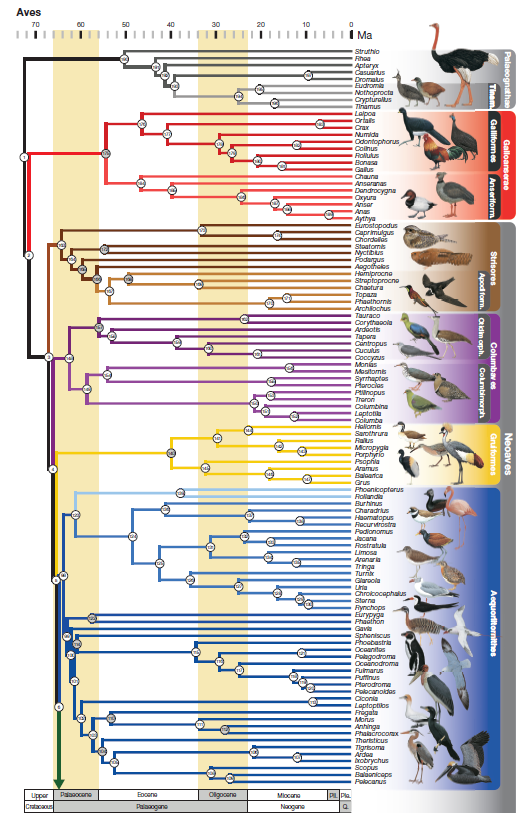
Figure 3a. First part of Bayesian phylogeny
from Prum et al. (2015).
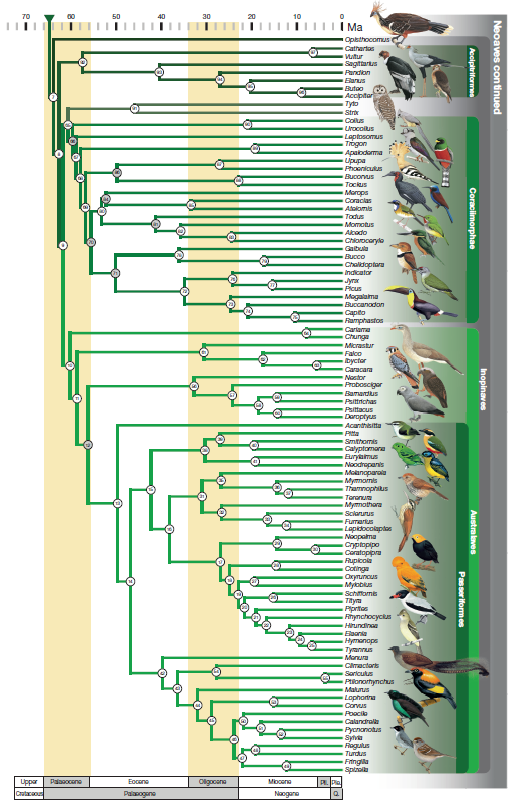
Figure 3b. Second part of Bayesian phylogeny
from Prum et al. (2015).
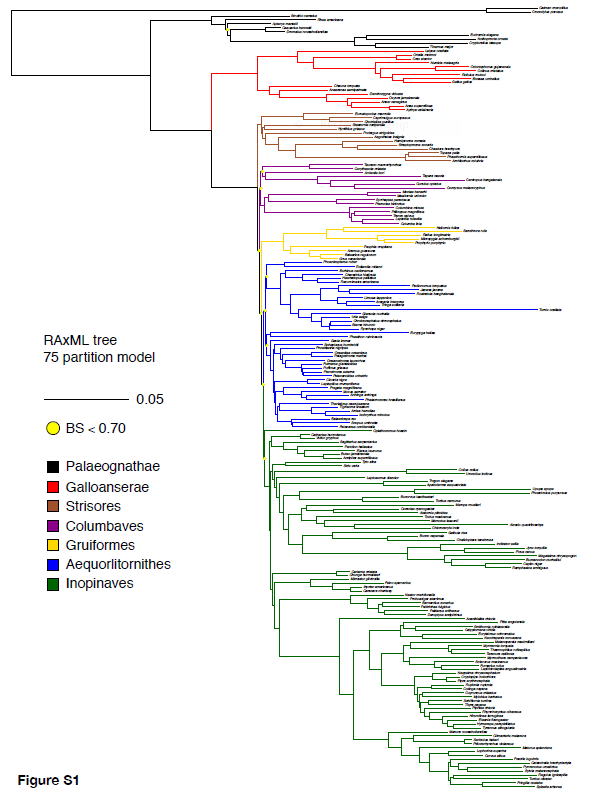
Figure
4. Maximum likelihood phylogeny in the supplemental material from Prum et al.
(2015). Yellow dots indicate nodes with <70% bootstrap support.
As
noted by Suh et al. (2015), the diversification of Neoaves seems to consist
largely of three radiations: a poorly resolved initial radiation at the base of
the Neoaves (consisting of Podicipediformes, Phoenicopteriformes,
Columbiformes, Pterocliformes, Mesitornithiformes, Cuculiformes,
Musophagiformes, Otidiformes, Caprimulgiformes, Apodiformes, Opisthocomiformes,
Gruiformes, Charadriiformes, and perhaps Eurypygiformes and Phaethontiformes,
although these latter two are likely the sister group to the core waterbirds)
followed by the better resolved radiations of the core waterbirds and core
landbirds. Thus, we have relatively strong support for the makeup and placement
of the core waterbirds and landbirds, but only poor to moderate support for the
placement of most other avian groups.
The
following motions propose to change the SACC higher-level linear sequence based
on the new phylogenetic studies, relying more heavily on the most data-rich
studies, those of Jarvis et al. (2014) and Prum et al. (2015). Subproposals
(a)-(b) would place together in the linear sequence all orders that constitute
the core landbirds. Subproposal (c) would do the same for the core waterbirds,
and place the Eurypygiformes and Phaethontiformes so as to precede the core
waterbirds. Subproposal (d) is a
bookkeeping change that should have been made some time ago. Subproposals
(e)-(h) would: (1) place together in the linear sequence all other orders that
appear to constitute the initial radiation of Neoaves, and (2) place these
orders in a linear sequence that seems to best represent the bulk of the
evidence. Given the uncertainty at the base of the tree, the placements of
these latter orders differ somewhat in the various studies, but many of these
different placements can be accommodated in a linear sequence.
An
appendix detailing the current SACC linear sequence, the linear sequence that
would result from approval of proposals (a)-(d), the linear sequence that would
result from approval of all proposals, and the linear sequence that would
result from following the Bayesian tree in Prum et al. (2015) appears at the end
of this proposal.
(a) Move Strigiformes
to precede Trogoniformes, and (b) move Cathartiformes + Accipitriformes to
precede Strigiformes.
One
of the most striking results of the recent DNA sequence studies, such as the
intron-based study of 169 species (Hackett et al 2008), the whole-genome study
of 48 species (Jarvis et al 2014), and the target-enrichment study of 198
species (Prum et al. 2015), and not found in Sibley and Ahlquist, was the lack
of a sister relationship between the falcons and caracaras on the one hand and
the hawks, eagles, kites, and vultures on the other. SACC recently removed the
Falconiformes sensu stricto from the
Accipitriformes and placed them towards the end of the linear sequence as
sister to a Passeriformes/Psittaciformes clade, but the
Cathartiformes+Accipitriformes were left in their former position between the
core waterbirds and the Gruiformes sensu
lato. Likewise, SACC maintained the Strigiformes in their traditional
placement between the Cuculiformes and the Caprimulgiformes.
These
new moves would reflect the phylogenetic relationships of the core landbird
clade mentioned above and would result in the following sequence at the end of
the classification: Cathartiformes, Accipitriformes, Strigiformes,
Trogoniformes, Coraciiformes, Galbuliformes, Piciformes, Cariamiformes,
Falconiformes, Psittaciformes, and Passeriformes. This sequence is in agreement
with the phylogenetic trees in Hackett et al. (2008), Jarvis et al. (2014), and
Prum et al. (2015), among others.
According
to Hackett et al. (2008), the Strigiformes are sister to the Coliiformes, and
this clade is sister to a clade consisting of the Leptosomiformes,
Trogoniformes, Upupiformes, Coraciiformes, and Piciformes. The Accipitriformes sensu lato (incl. Cathartidae) are
sister to this entire clade. Although these results received poor bootstrap
support (< 50%), all of the more recent studies support this result, some
with strong support.
For
example, Jarvis et al. (2014), in their whole genome study, recovered a clade
consisting of the Coliiformes, Leptosomiformes, Trogoniformes, Bucerotiformes,
Coraciiformes, and Piciformes (Coraciimorphae, sensu Jarvis et al.), with the Strigiformes sister to this clade,
and the Accipitriformes sensu lato sister
to this larger clade. Bootstrap support for the entire clade (“Afroaves” sensu Jarvis et al.) was 100%, support
for the Accipitriformes as sister to all other taxa was 100%, and support for
the Strigiformes as sister to the rest (except for the Accipitriformes) was
84%. Prum et al. (2015) also recovered this arrangement, except that the
Accipitriformes were sister to a clade consisting of the Strigiformes and
Coraciimorphae + Cariamiformes, Falconiformes, Psittaciformes, and
Passeriformes (these four orders constituting the Australaves), making
“Afroaves” paraphyletic. Posterior probabilities for their results were all 1.0
and ML bootstraps were all >70% (finer resolution was not provided for the
bootstraps).
The
current SACC linear sequence has a monophyletic Coraciimorphae but, as noted
above, places the Cathartiformes+Accipitriformes far from the Coraciimorphae
and also separates the Strigiformes from these two groups. By moving the
Strigiformes to precede the Trogoniformes, and then moving the
Cathartiformes+Accipitriformes to precede the Strigiformes, SACC would achieve
the following: (1) make the core landbird clade monophyletic; (2) place the
Strigiformes in the correct position as sister to the Coraciimorphae; and (3)
position Cathartiformes+Accipitriformes in the linear sequence to reflect a
sister relationship with the rest of “Afroaves” (as in Jarvis et al. 2014) or
to reflect a sister relationship with the
Strigiformes+Coraciimorphae+Australaves clade (as in Prum et al. 2015).
(c) move Eurypygiformes
and Phaethontiformes to precede Sphenisciformes.
Numerous
studies have identified a core waterbird clade consisting of the Gaviiformes,
Procellariiformes, Sphenisciformes, Pelecaniformes, Suliformes, and
Ciconiiformes, but not the Phaethontiformes, which are currently embedded
within the waterbird clade in the SACC linear sequence. Sibley and Ahlquist
(1990), using DNA-DNA hybridization, found that their representatives of these
orders formed a monophyletic group, that this group of waterbirds was embedded
deep within the Neoaves, that the Phaethontiformes were the sister group to
this waterbird clade, and that within the clade the Gaviiformes and
Procellariiformes were sister groups. Hackett et al. (2008) also found that the
Gaviiformes, Procellariiformes, Sphenisciformes, Pelecaniformes, Suliformes,
and Ciconiiformes formed a clade, with reasonably strong support (89%
bootstrap), and that this clade was embedded within the Neoaves. However, the
Gaviiformes were sister to the rest of this group (94% bootstrap).
The
presence of this core waterbird clade has been remarkably consistent in the
recent studies. The UCE-based study of McCormack et al. (2013) found strong
support for the waterbird clade, and Kimball et al. (2013) also recovered this
clade. Jarvis et al. (2014) found that the Gaviiformes, Procellariiformes,
Sphenisciformes, Pelecaniformes, and Suliformes formed a clade, and presumably
the Ciconiiformes would have formed part of this group had a representative
been included in the study. Burleigh et al. (2015) and Prum et al. (2015) found
a waterbird clade of the same composition but including Ciconiiformes, with
moderate to strong support: 73% bootstrap in Burleigh et al., 1.0 pp and
>70% bootstrap in Prum et al.
Three of the recent studies provided support
for Sibley’s result of the Phaethontiformes as sister to the waterbird clade.
In Jarvis et al. (2014) and Prum et al. (2015), the Phaethontiformes and the
Eurypygiformes formed a clade that was sister to the waterbird clade, with
moderate to strong support (70% bootstrap in Jarvis et al., 1.0 pp and >70%
bootstrap in Prum et al.), and this clade was also sister to the waterbird
clade in at least one of the trees in Kimball et al. (2013), although with poor
support.
The Eurypygiformes have generally been
difficult to place, and the results of Jarvis et al. and Prum et al. provide
some of the only reasonable support for placement of this order. Hackett et al.
(2008), for example, had them as sister to the Caprimulgiformes-Apodiformes
clade, but support was not strong (61% bootstrap). Several trees in Kimball et
al. (2013) placed them as sister to the Phaethontiformes, as did McCormack et
al. (2013), although their placement beyond this was uncertain.
All studies provide strong support for the
monophyly of the Gaviiformes, Procellariiformes, Sphenisciformes,
Pelecaniformes, Suliformes, and Ciconiiformes as a “core waterbirds” group,
with reasonable support for the Phaethontiformes and Eurypygiformes as sisters
to this group. The current SACC sequence places the Sphenisciformes directly
after the Podicipediformes and Phoenicopteriformes. Next in the linear sequence
are the Procellariiformes, Phaethontiformes, Ciconiiformes, Suliformes, and
Pelecaniformes; this is the core waterbird group identified by the genetic
studies except for the presence of the Phaethontiformes. By moving the
Eurypygiformes and Phaethontiformes to precede the Sphenisciformes, SACC would
achieve the following: (1) make the core waterbird group monophyletic, and (2)
place Eurypygiformes and Phaethontiformes in the correct position as indicated
by Jarvis et al. and Prum et al., as sister group to the core waterbirds.
(d) Reverse the linear
sequence of Podicipediformes and Phoenicopteriformes.
SACC
has long recognized Podicipediformes + Phoenicopteriformes as a clade, but for
some reason the order with more genera and species (Podicipediformes = 6
genera, 22 species) precedes the one with fewer genera and species
(Phoenicopteriformes = 1 genus, 6 species) in the classification, in contradiction
to the conventions for linear sequencing. This subproposal corrects this error.
(e) Move Columbiformes
to follow Podicipediformes, and (f) move Cuculiformes, Caprimulgiformes and
Apodiformes to follow Columbiformes.
In
the current SACC sequence, the Columbiformes
and Cuculiformes (with Opisthocomiformes, see below, in between) precede the
Strigiformes, and the Caprimulgiformes and Apodiformes follow the Strigiformes.
If subproposals (a) and (h) pass, these groups will occur together in the
linear sequence.
Hackett
et al. (2008), Jarvis et al. (2014), and Burleigh et al. (2015) found close
relationships among the Columbiformes, Pterocliformes, Mesitornithiformes,
Phoenicopteriformes, and Podicipediformes, Jarvis et al. with strong support.
The relevant discrepancy in their respective results was the addition of the
Phaethontiformes to the clade in Hackett et al (2008) and Burleigh et al.
(2015), although with poor support. The position of the Phaethontiformes well
outside of this clade (as or in the sister group to the core waterbird clade)
in the studies of Sibley and Ahlquist (1990), Kimball et al. (2013), Jarvis et
al (2014), and Prum et al. (2015) along with the poor support for this clade in
Hackett et al. (2008) and Burleigh et al. (2015), suggests that the placement
as sister to the waterbirds, as above in (c), is likely more appropriate.
Other
orders that have consistently shown up near the base of the Neoaves are the
Cuculiformes, Caprimulgiformes, and Apodiformes. In Jarvis et al. (2014), representatives
of these orders were placed in the clade that is sister to all other Passerea
(which encompasses all Neoaves except for their Columbea), with reasonably
strong bootstrap support (91%). In Hackett et al (2008), the Caprimulgiformes
formed part of the clade that includes most of the orders in the Columbea of
Jarvis et al., mirroring in part the Metaves of Fain and Houde (2004), although
bootstrap support was weak (< 50%). The Cuculiformes, Gruiformes, and
Otidiformes formed a clade that was sister to the Musophagiformes + the core
waterbird clade – a result that received reasonable support (81%) although this
support was much reduced (<50%) in the extended dataset of Kimball et al.
(2013). There was little support for relationships of these orders in McCormack
et al. (2013) and Burleigh et al. (2015). Prum et al. (2015) found the
Caprimulgiformes + Apodiformes to be sister to all other Neoaves, with their
Columbaves (including Cuculiformes, Otidiformes, Musophagiformes,
Columbiformes, Pterocliformes, and Mesitornithiformes) the next successive
sister, thus reversing the linear sequence proposed here.
By
moving the Pterocliformes and Columbiformes to follow the Podicipediformes,
SACC would achieve the following: (1) place them in linear sequence to reflect
a monophyletic “Columbea” sensu Jarvis
et al., a group that also has strong support (although with the addition of
Phaethontidae) in Hackett et al.; and (2) place the Columbea in linear sequence
directly following the Galloanseres, in keeping with their position as sister
to the rest of Neoaves (Jarvis et al. 2014), a position also consistent with
that in Hackett et al., although at odds with Prum et al.
By
moving the Cuculiformes, Caprimulgiformes and Apodiformes to follow the
Columbiformes (and Columbimorphae), SACC would position these orders to: (1)
reflect their sister relationship (along with the extralimital Musophagiformes
and Otidiformes) with the rest of the Passerea (according to Jarvis et al
2014), and (2) reflect their position as successive sisters (although in
reverse order) at or near the base of the Neoaves (as in Prum et al. 2015).
(g) Move the Gruiformes
and Charadriiformes to precede the Eurypygiformes.
These
are groups for which many of the phylogenies rather profoundly disagree; this
motion generally follows the results of Jarvis et al. (2014) and Prum et al.
(2015). Jarvis et al. (2014) placed the Charadriiformes and Gruiformes (along
with the hoatzin; see below) as sisters to a clade consisting of the core
waterbirds + Phaethontiformes + Eurypygiformes + the core landbirds. Prum et
al. (2015) placed the Charadriiformes (along with the Phoenicopteriformes +
Podicipediformes) as sister to the core waterbird clade + Phaethontiformes +
Eurypygiformes, and the Gruiformes as sister to this large clade.
(h) Move the
Opisthocomiformes to precede the Gruiformes.
The
position of the Opisthocomiformes on the avian tree has long been unsettled. Opisthocomus has been placed near the
Galliformes, Cuculiformes, Musophagiformes, Columbiformes, and other groups.
Unfortunately, and perhaps unsurprisingly, recent phylogenetic and phylogenomic
studies have not resolved the placement of this group. In Hackett et al.
(2008), Opisthocomus was sister to a
group containing the core waterbirds, Musophagiformes, Gruiformes,
Cuculiformes, and Otidiformes, but bootstrap support was weak. In Jarvis et al.
(2014), Opisthocomus was sister to
the Gruiformes + Charadriiformes (bootstrap = 91%), and this group was sister
to the core waterbirds, Phaethontiformes, Eurypygiformes, and core landbirds.
In Prum et al. (2015), in contrast, Opisthocomus
was sister to the landbirds, with strong bootstrap support in the published
Bayesian tree but no support in the tree in the supplementary material.
The
current SACC linear sequence places the Opisthocomiformes between the
Columbiformes and Cuculiformes, breaking up groups that are either part of
sister groups (Prum et al. 2015) or that form part of successive sister groups
to the rest of Neoaves (Jarvis et al. 2014), and that should be placed close
together in the linear sequence following either study. Placing Opisthocomus according to Prum et al.
(2015) would move it to a position between the core waterbirds and core
landbirds, very different from its current placement, whereas following Jarvis
et al. (2014) would move it only slightly (see Appendix). I propose that the
Opisthocomiformes be placed between the Apodiformes and the Gruiformes,
essentially following the latter – this is something of a middle ground between
the current placement and that of Prum et al. (2015).
Recommendation:
I
strongly recommend voting in favor of some of these proposals (a-d), and I
endorse the others as well. A new linear sequence that includes some or all of
these changes would reflect the best data on higher-level avian relationships
much better than does the current SACC sequence. The alternative would be to
wait until phylogenomic studies with better taxon sampling and tree-wide
consistent support are available, but consistent resolution of the initial
radiation of Neoaves may be difficult to achieve, and I would say that some of
these changes need to be made now.
If
SACC approves only (a)-(d), then they would have to decide whether the
remaining orders fit in their “new” positions (see the appendix below).
Approval of all proposals (a)-(h) would generally follow the bulk of the
phylogenetic evidence currently available. An alternative would be to follow
the published tree of the most recent major study, that of Prum et al. (2015).
Their published Bayesian tree received strong support at almost all nodes, but
most of the deeper nodes were poorly supported in the maximum likelihood tree
in their supplementary material. The differences between the (a)-(h) linear
sequence and that derived from Prum et al. are in the placement of the
Phoenicopteriformes + Podicipediformes (early in the a-h sequence, somewhat
later in Prum et al.), the relative placement of the Pterocliformes +
Columbiformes, Cuculiformes, and Caprimulgiformes + Apodiformes (in this
sequence in (a)-(h), in the sequence Caprimulgiformes + Apodiformes,
Cuculiformes, and Pterocliformes + Columbiformes in Prum et al.), and the
placement of the Opisthocomiformes (preceding Gruiformes in (a)-(h), between
the core waterbirds and core landbirds in Prum et al.). Prum et al. placed all
of the “initial radiation waterbirds” together preceding the core waterbirds,
and this has some intuitive appeal in that all the waterbirds are together in
the linear sequence. However, the (a)-(h) sequence causes less disruption to
our current linear sequence, leaving Phoenicopteriformes + Podicipediformes
near the beginning of the sequence, and leaving intact the linear sequence of
Columbiformes, Cuculiformes, Caprimulgiformes + Apodiformes (although in the
current sequence, Opisthocomiformes and Strigiformes are embedded among these
orders). To me, this seems like the slightly better course until we get more
definitive data on the initial Neoavian radiation.
A
slightly different version of this proposal (lacking discussion of
Sphenisciformes and Opisthocomiformes but including discussion of Gaviiformes
and Pterocliformes) formed NACC Proposal 2016-C-6. This proposal passed unanimously.
Literature
Cited:
Burleigh, J. G., R. T. Kimball, and E.
L. Braun. 2015. Building the avian tree of life using a large-scale, sparse
supermatrix. Mol. Phylogenet. Evol. 84: 53-63.
Fain, M. G., and P. Houde. 2004.
Parallel radiations in the primary clades of birds. Evolution 58: 2558-2573.
Groth, J. G., and G. F. Barrowclough.
1999. Basal divergences in birds and the phylogenetic utility of the nuclear
RAG-1 gene. Mol. Phylogenet. Evol. 12: 115–123.
Hackett, S. J., et al. 2008. A
phylogenomic study of birds reveals their evolutionary history. Science 320:
1763-1768.
Jarvis, E. D., et al. 2014. Whole-genome
analyses resolve early branches in the tree of life of modern birds. Science
346: 1320-1331.
Kimball et al. 2013. Identifying local
biases in large datasets: a case study using the avian tree of life. Mol.
Phylogenet. Evol. 69: 1021-1032.
McCormack et al. 2013. A phylogeny of
birds based on over 1,500 loci collected by target enrichment and
high-throughput sequencing. PLoS ONE 8: e54858.
Prum, R. O., et al. 2015. A
comprehensive phylogeny of birds (Aves) using targeted next-generation DNA
sequencing. Nature 526: 569–573.
Sibley, C. G., and J. E. Ahlquist. 1990.
Phylogeny and classification of birds: a study in molecular evolution. Yale
Univ. Press, New Haven, CT.
Suh, A., L. Smeds, and H. Ellegren.
2015. The dynamics of incomplete lineage sorting across the ancient adaptive
radiation of Neoavian birds. PLOS Biology 13(8): e1002224, 18 pp.
Terry Chesser, October 2016
Appendix: The current SACC sequence of orders, the
linear sequence that would result from approval of proposals a-d above, the
linear sequence that would result from approval of all proposals above, and the
linear sequence that would result from following the Bayesian tree in Prum et
al. (2015). Orders that form part of the poorly resolved initial radiation of
Neoaves are in bold, those in the core waterbird clade are highlighted in blue,
and those in the core landbird clade are highlighted in green.