Proposal
(752) to South American Classification Committee
Split Sclerurus mexicanus into multiple
species
The following proposal is a set of
hierarchical taxonomic schemes for the Sclerurus
mexicanus complex. It expands upon Proposal 603 with newly published vocal
analyses (Cooper & Cuervo 2017). This proposal is meant to be considered
sequentially (i.e., Part I must be approved to consider Part II).
Part I
Split Sclerurus mexicanus into two species: Sclerurus mexicanus and Sclerurus obscurior.
Effect on SACC: Sclerurus mexicanus would be split into two species that are
assumed to be parapatric in the Darién Gap. All known South American
populations would become S. obscurior;
S. mexicanus would be moved to the
hypothetical list.
Background: The genus Sclerurus currently contains six widespread, polytypic species. Sclerurus mexicanus has the broadest
distribution of any Sclerurus, with
seven subspecies occurring from northern Mexico to southern Brazil (Cooper and
Barragan 2017; Fig. 1). Despite the recognized diversity and broad distribution,
differences are minimal (Remsen 2003), and subspecies distributions are still
incompletely known (Cooper and Barragan 2016; Cooper and Cuervo 2017). d’Horta
et al. (2013) found that Sclerurus
mexicanus is not monophyletic as currently defined. Rather, three major
clades were discovered that form a polytomy in the rufous-throated leaftosser
clade: (1) S. mexicanus (subspecies mexicanus and pullus) in North America; (2) S.
rufigularis; and (3) S. mexicanus
(subspecies obscurior, andinus, macconnelli, and peruvianus)
in South America. Similarly, recent vocal analyses only found S. rufigularis to be statistically
diagnosable when analyses allowed for multiple species within the S. mexicanus complex (Cooper and Cuervo
2017).
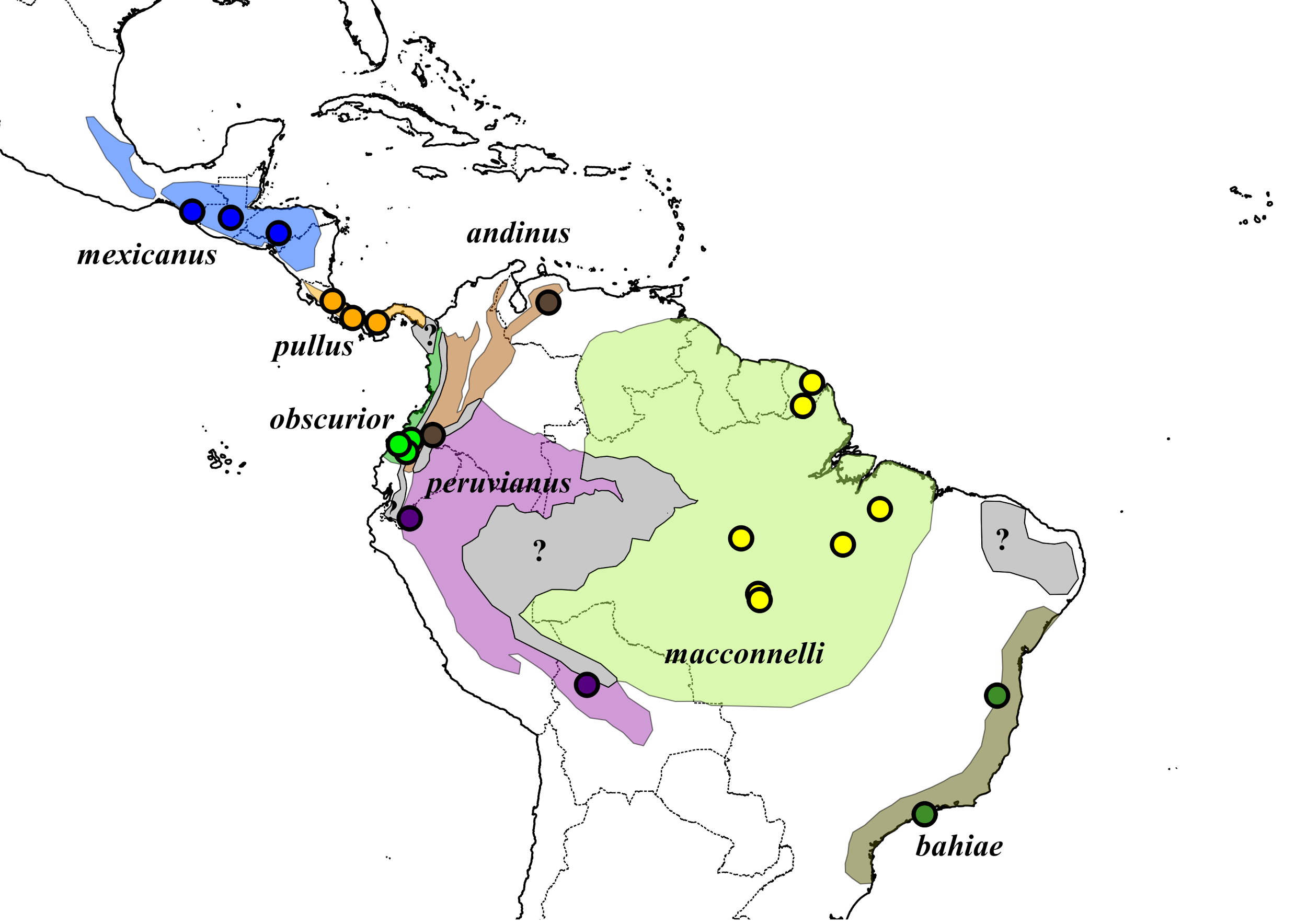
Figure 1. A map of
subspecific distributions within Sclerurus
mexicanus. Points represent localities from which vocalizations were
sampled. Figure from Cooper and Cuervo (2017).
Within the S.
mexicanus group, North and South American populations appear wholly
allopatric, diagnosably monophyletic (d’Horta et al. 2013), and vocally
distinct (Cooper and Cuervo 2017). Unless all rufous-throated leaftossers are
considered a single species (including S.
rufigularis), the most prudent decision is to split S. mexicanus into two taxa with no known geographic overlap:
1.
Sclerurus mexicanus (Sclater 1856). Suggested English name:
Central American Leaftosser. Type locality: Cordoba, Veracruz, Mexico. This
species contains two subspecies: mexicanus
(from northern Nicaragua north to east Mexico) and pullus (from northern Costa Rica south to southern Panama). This
includes certus (Chubb 1919, from
Guatemala), which is considered a synonym of mexicanus (Hellmayr 1925).
2.
Sclerurus obscurior (Hartert 1901). Suggested English name: Dusky
Leaftosser. Type locality: Lita, Esmeralda, Ecuador (ca. 600m). This species
contains five subspecies: obscurior
(the Choco lowlands), andinus (the
mid-montane Andes of Colombia, Ecuador, and Venezuela), peruvianus (the eastern foothills of the Andes and western Amazonia
in Peru, Ecuador, Colombia, and probably Brazil), macconnelli (eastern Amazonia and the Guianan shield), and bahiae (the Atlantic Forest of Brazil).
Thus, if accepted, all confirmed populations
in South America would become subspecies of S.
obscurior.
Part IIA
Split Sclerurus mexicanus into Sclerurus mexicanus and Sclerurus pullus. Advisory only to NACC
Effect on SACC: This would only become
important if the population pullus is
confirmed in Northern Colombia. Sclerurus
pullus is known from adjacent eastern Panama. This would have no effect on
the main SACC list at the present time; S.
mexicanus would be removed from the hypothetical list and replaced with S. pullus.
Background: Sclerurus mexicanus and Sclerurus
obscurior have no known regions of overlap, with no confirmed records of Sclerurus mexicanus (sensu Part I) in
the SACC region. However, given that S.
mexicanus pullus occurs in eastern Panama immediately adjacent to Colombia,
it is possible that the taxon will be found in the SACC area in the future.
Despite limited vocal data, differences were recovered between S. m. mexicanus and S. m. pullus (Cooper and Cuervo 2017) and populations are
reciprocally monophyletic (d’Horta et al. 2013). A split of these two taxa is
therefore warranted. This will have no effect at the present time on the SACC
list, but will change any future records of S.
mexicanus from northern Colombia to S.
pullus, as follows:
Sclerurus pullus (Bangs 1902).
Suggested English name: Isthmian Leaftosser (thus altering S. mexicanus to Tawny-throated or Mexican Leaftosser). Type
locality: Boquete, Panama. Distributed from northern Costa Rica through the
Darién in Eastern Panama. This species may occur in the Darién and Urabá
regions of Colombia, and may exist parapatrically with S. obscurior. This species includes the synonym anomalus (Bangs and Barbour 1922), which
has erroneously been synonymized with andinus
in the past (Peters 1951).
Part IIB
Split Sclerurus
obscurior into three species: Sclerurus
obscurior, Sclerurus andinus, and
Sclerurus macconnelli.
Effect on SACC: Sclerurus obscurior would become three allopatric species, with Sclerurus obscurior being retained by
lowland Choco populations. Andean populations would become Sclerurus andinus, and Amazonian/Atlantic Forest populations would
become Sclerurus macconnelli.
Background: d’Horta et al. (2013) recovered
multiple monophyletic clades within South American S. obscurior (sensu Part I). These groups correspond to all
subspecies for which genetic data were available (i.e. all subspecies except bahiae), with the shortest branch lengths
for the division between S. o. peruvianus
and S. o. macconnelli (Fig. 2).
Given this short branch length, it is reasonable to consider S. peruvianus and S. macconnelli conspecific (but see Part III below). Cooper and
Cuervo (2017) tested multiple different partitioning schemes on Sclerurus songs and found the highest
support for a similarly conservative partitioning scheme that splits obscurior and andinus but retains peruvianus,
macconnelli, and bahiae as a single polytypic species.
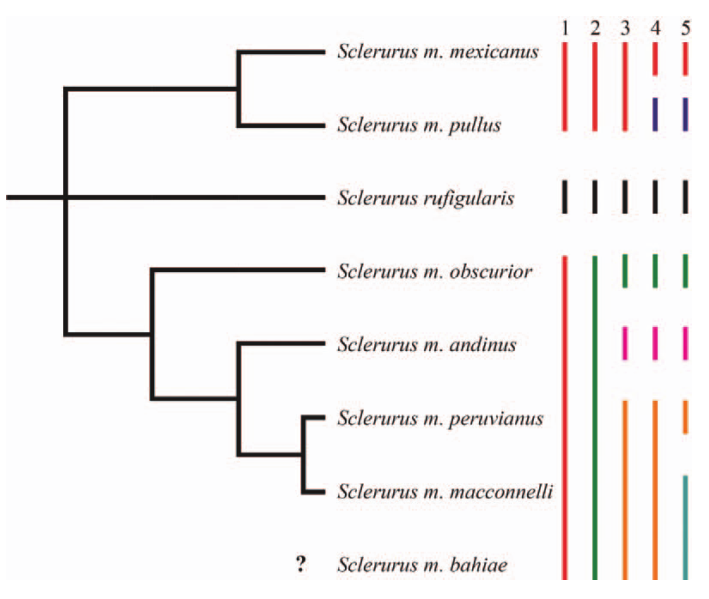
Figure 2. Species partitioning schemes tested
for the S. mexicanus complex. Branch
lengths are not to scale, but are presented proportionally and the tree depicts
monophyletic assemblages recovered by d’Horta et al. (2013). Colored lines,
labeled 1-5, represent groupings discussed in Cooper & Cuervo (2017); with
respect to this proposal, partition 2 corresponds to Part I, partition 4 to
Part II, and partition 5 to Part III. Figure from Cooper and Cuervo (2017).
Genetic data supports the split of andinus from obscurior (d’Horta et al 2013): “Along the continuum of humid
forests from the Chocó lowlands to the slopes of the western Andes, two
lineages appear segregated elevationally: S.
obscurior, and S. andinus found
locally from about 1000 m (often up to 2000 m). The two lineages are
potentially syntopic at an intermediate point of the elevational and ecological
gradient, where no obvious physical barrier is in place… the lowland Chocó
(i.e. S. obscurior) and the Andean
foothill species (i.e. S. andinus)…[last]
shared a common ancestor in the Early Pliocene, between 3.6 and 6.0 Ma”.
Based on these data, Part IIB therefore
proposes elevation of S. andinus and S. macconnelli to species level. S. obscurior would then become a
monotypic taxon restricted to the Choco (with the suggested English name ‘Dusky
Leaftosser’). The descriptions for these new taxa would be as follows:
1.
Sclerurus andinus (Chapman 1914). Suggested English name:
Andean Leaftosser. Type locality: Buenavista, above Villavicencio, Colombia, on
the E slope of the Eastern Andes (ca. 1370 m). This subspecies resides in humid
submontane forest in western Venezuela (the main Andes and the Sierra de
Perijá), all three Andean ranges of Colombia, and in Western Ecuador. Records
from northeastern Ecuador and southwestern Ecuador (El Oro) may refer to this
taxon.
2.
Sclerurus macconnelli (Chubb 1919). Suggested English name:
Long-billed Leaftosser (as opposed to Short-billed Leaftosser S. rufigularis with which this species
is sympatric; MacConnell’s, Amazonian, or Guianan Leaftosser [as in Cory &
Hellmayr] are also possible English names). Type locality: Ituribisci River,
Guyana. This species would consist of three subspecies: peruvianus of the eastern foothills of the Andes and western
Amazonia; macconnelli of central and
eastern Amazonia and the Guianan shield; and bahiae of the Atlantic Forest of Brazil. Records from Ceará,
halfway between the distributions of macconnelli
and bahiae, are best left
unidentified at this time. While parapatry is assumed between peruvianus and macconnelli, the contact zone is not well known.
Part III
Split Sclerurus
macconnelli into two species: Sclerurus
macconnelli and Sclerurus peruvianus.
Effect on SACC: Amazonian populations of S. macconnelli would be split into two
species, with peruvianus occupying
the western basin and Andean foothills and macconnelli
occupying the lower basin, the Guianan shield, and the Atlantic Forest.
Background: While defined vocal differences
between S. macconnelli subspecies
(sensu Part IIB) were not recovered (Cooper & Cuervo 2017), d’Horta et al.
(2013) recovered two monophyletic groups aligning to the described populations
of peruvianus and macconnelli. Per d’Horta et al. (2013):
“... S. macconnelli and S. peruvianus … are in close
geographical proximity in southern Peru and Bolivia but seem to occupy
different elevations along the cis-Andean foothills”. The magnitude of the
differentiation between macconnelli
and peruvianus is less than any other
branching event within S. mexicanus
sensu lato, including the relationship between S. mexicanus and S. pullus.
Regrettably, no genetic data is presently
available for bahiae, and its
relationship to the rest of the S.
macconnelli complex is uncertain. Thus, Part III opts for the elevation of S. peruvianus to species level while
retaining bahiae as a subspecies of
the geographically proximate S.
macconnelli.
S. peruvianus (Chubb 1919).
Suggested English name: Peruvian Leaftosser (as in Cory & Hellmayr). Type
locality: Yurimaguas, Loreto, Peru. This species replaces macconnelli in northwestern Amazonia and in the higher elevations
of the Andean foothills in southern Peru and Bolivia. Exact range limits are
unknown, but it is known to occur on both sides of the Napo/Amazon Rivers in
Ecuador and Colombia.
Recommendation
Given the amount of genetic and vocal data
available, we recommend the acceptance of Part I and Part II (both A and B).
Five species are repeatedly delimited using both vocal and genetic data, and
best represent the diversity within S.
mexicanus sensu lato. These decisions would remove S. mexicanus from the SACC list and replace it with three species: S. obscurior, S. andinus, and S.
macconnelli. One additional species should be considered hypothetical
within the SACC area (S. pullus)
pending further surveys of the Darién Gap.
Given the present data, we recommend holding
off on Part III. The relationship of bahiae
to peruvianus and macconnelli is not resolved, and there is
no reliable way to identify these taxa in the field at the present time short
of collecting vouchers or genetic samples.
Literature cited
Cooper, J. C. & A.
M. Cuervo. 2017. Vocal variation and species limits in the Sclerurus mexicanus complex. The
Wilson Journal of Ornithology 129:13-24.
Cooper, J. C. &
Barragán, D. 2017. Tawny-throated Leaftosser Sclerurus mexicanus. Neotropical Birds Online
<http://neotropical.birds.cornell.edu/>.
d’Horta, F. M., A. M.
Cuervo, C. C. Ribas, R. T. Brumfield & C. Y. Miyaki. 2013. Phylogeny and
comparative phylogeography of Sclerurus (Aves:
Furnariidae) reveal constant and cryptic diversification in an old radiation of
rain forest understory specialists. Journal
of Biogeography 40:37-49.
Other references in SACC bibliography.
Jacob C. Cooper & Andres M. Cuervo, May 2017
========================================================
Comments
from Jaramillo:
“Part I – YES, data look solid to me, and include multiple independent sets of
data. Note that Central American Leaftosser does not work for me.
“Part
IIA – YES, assuming it eventually is found in Colombia.
“Part
IIB – YES, I find it powerful that obscurior
and andinus are syntopic, and replace
each other elevationally. It is unfortunate that bahiae was not sampled molecularly, as it may shift the tree
perhaps?
Part
III – NO, unresolved as noted in the proposal. Data needed for bahiae, as well as no reliable way to
identify taxa in field!”
Comments
from Stiles:
"YES to part 1. YES to part 2. The split between mexicanus and pullus is deep, suggesting isolation across the Nicaraguan gap in
mountain habitats. YES to part 3,
given the genetic and vocal data and the likelihood of parapatry (or even
sympatry?) at intermediate elevations. NO
to part 4: data are not sufficient for this split at this time."
Comments
from Claramunt:
" NO. Quoting Cooper & Cuervo (2017:13): “…revising
species limits in the group is complicated by subtle phenotypic variation
between lineages and an incomplete understanding of the limits of their distribution.” I fully agree
and this exactly what is missing in this proposal: a better understanding of
phenotypic geographic variation. We know that there is important geographic
variation in plumage (Remsen 2003), mitochondrial DNA (d’Horta et al. 2013),
and vocalizations (Cooper & Cuervo 2017). What we don’t know yet is whether
that variation is just geographic variation in a widespread species or
indicative of multiple species. In particular, a strong match between genetic
and phenotypic subdivisions would suggest reproductive isolation in
differentiated lineages, thus species status. What we observe here is a general
correlation between the different sets of data, but I don’t see evidence of a
strong match across traits.
"For example, affinities of the birds from E Panama are
uncertain; plumage suggests affinities with the South American clade: birds
there have been referred to obscurior and andinus (Wetmore1972,
Remsen 2003). On the other hand, the bird included in d’Horta et al. from E
Panama carried a Central American haplotype (it is identified as pullus in
the paper, but I don’t know if that was based on plumage or a posteriori,
based on DNA). Songs from this region were not analyzed by Cooper & Cuervo
(2017), but a song from this region in xeno-canto sounds to me like a perfect
intermediate between those of Central America and NW South America. So, right
now, we don’t know if this pattern reflects primary intergradation of traits, a
hybrid zone, or sympatry of two isolated lineages.
"Similarly, birds from the Pantepui region have been assigned
to andinus but carry a macconnelli mtDNA, and birds from W
Brazilian Amazonia have been referred to peruvianus but carry macconnelli
mtDNA. Thus, it seems that there are broad mismatches between mitochondrial and
plumage variation, thus suggesting that we are seeing the vagaries of
polymorphic traits within a large and geographically structured population,
rather than discrete lineages evolving independently. A detailed analysis of
plumage variation would be very informative.
"Song variation is also complex and, aside from a general
large-scale correspondence, I don’t see evidence that it matches either plumage
or mtDNA, and the statistical design used by Cooper & Cuervo is problematic
in several respects. First, they did not analyze songs from both sides of
putative contact zones, like Darién Chocó (pullus-obscurior), the W
foothills of the W Andes (obscurior-andinus, all songs of andinus
are from the E slopes of E cordilleras) or Amazonian Andes forelands (peruvianus-macconnelli).
Therefore, the possibility of finding intermediate songs and mismatches was
minimized. Second, univariate pairwise comparisons of song traits was
problematic for two reasons: 1) differences in group means do not prove that
groups are discrete units (cannot distinguish clines from discrete variation),
and 2) incurred in a serious problem of multiple testing (Appendix 2 contains
196 P-values; standard methods for adjusting P-values for multiple test result
in none of the pairwise tests being statistically significant! Note that none
of the P-values are particularly low). Therefore, no statistical evidence
there. The discriminant analysis does provide some support for one of the
proposed taxonomies, but again, the lack of samples near putative contact zones
may have biased the analysis towards good levels of discrimination. Finally,
from the perspective of the biological species concept, there is no information
regarding the potential effect of the geographic variation in song on
reproductive isolation. Actually, birds seem to respond to songs from distantly
related subspecies and they can sometimes switch to a song that is more similar
to that of other subspecies (Cooper & Cuervo 2017:16); this kind of
information is usually taken as evidence of conspecificity under the biological
species concept.
"Finally, the phylogeny of d’Horta et al. (2013) raises the
possibility that mexicanus is not monophyletic, but the evidence is weak
(no statistical support either way). In sum, I can’t find the evidence for the
existence of multiple species within mexicanus."
Comments from Zimmer:
“YES. I think that the genetic and vocal
differences of all Central American populations north and west from the
highlands of western Panama from those of South America provide strong evidence
for at least a two-way split in this complex.
Part IIA. Split Sclerurus mexicanus into S. mexicanus and S. pullus. YES
(tentatively). Again, I think that the
genetic differences are persuasive, and a split here would fit an established
biogeographic pattern of differentiation across the inter-montane gap between
Nicaragua and the Talamanca-Chiriqui highlands.
However, my YES vote comes with the caveat that the situation in the
Darién of eastern Panama really muddles the overall picture. The Proposal treats the birds of eastern
Panama as being referable to S. [m.]
pullus, but other authors (e.g. Remsen in HBW Volume 8) have characterized
the situation in eastern Panama as being one of elevational parapatry, with andinus inhabiting the lowlands, and obscurior replacing it in the
highlands. Looking at this from the
perspective of biogeography, the typical pattern of taxon-replacement that we
see in eastern Panama, is for Talamanca-Chiriqui highland birds to drop out in
the isolated mountains of central Panama (e.g. Coclé-Panama provincial border
region), and for lowland birds to extend eastward at least to the Bayano River
valley before being replaced in the lowlands of Darién by taxa typical of the
Chocó region of Colombia and northwestern Ecuador, with taxa occupying the
Darién highlands either unique to that region, or, showing affinities to Andean
taxa in Colombia. To me, the obvious
break in vocal characters among all “Tawny-throated Leaftossers” is between
Central American birds (verifiable east/south to the highlands of western
Panama) and South American birds east of the Andes. It doesn’t make much sense to me that the
leaftossers from eastern Panama would be referable to pullus. The scenario that
Van proposed in HBW (andinus in the
lowlands and obscurior in the
highlands) for eastern Panama makes more sense to me, and if accurate, the
apparent elevational parapatry and pattern of taxon replacement would argue for
splitting andinus and obscurior from one another, a split that
would probably be supported by plumage differences. I am not familiar with the voices of the
eastern Panamanian mexicanus, and it
doesn’t appear that Cooper & Cuervo (2017) included vocal samples from the
region in their vocal analysis, so I have no confidence that the distributional
boundaries of pullus, andinus and obscurior are being accurately
portrayed. Because of this, and taking
into consideration Santiago’s well-reasoned arguments regarding the lack of
clarity as regards phenotypic variation and congruence of the different data
sets, I’m not willing to go further in splitting up this complex until we know
more. I would echo Santiago’s calls for
better vocal sampling and analysis from both sides of putative contact
zones. So, for now at least, I’m a NO on
Part IIB, and Part III of this proposal.”
Comments
from Pacheco:
“NO. As stressed by Santiago and Kevin, I consider for the moment that data
(vocal, genetic) of some populations are still unavailable so that they can
have a more robust understanding of the specific limits in this complex.
“There
is no assurance that the birds of eastern Panama are "pullus". There is no definition about the affiliation of the
populations of the Pantepui (despite being the type-locality of macconnelli) and the Atlantic Forest. My opinion is that the geographic coverage and
the number of song samples is not enough to guarantee the proposed taxonomic
suggestions. More importantly, vocal analysis did not find perfect congruence
with genetics. Tentatively, I vote YES only for Part I.”
Comments
from Remsen:
“YES to all proposed splits. Although I
strongly appreciated the well-reasoned conservative comments of Santiago and
others, I am strongly influenced by handling specimens of all these taxa and by
the likely elevational separation of taxa that for me is a nail-in-coffin piece
of evidence for species rank. The
differences among S. mexicanus
populations sensu lato are greater
than those between Amazonian populations of S.
mexicanus and S. rufigularis. Continued maintenance of all of these taxa as
a single species masks a lot of what I would consider species-level
biodiversity. Philosophically, in this
group repairing and refining problems in species limits of all taxa recommended
as splits in the proposal (as in taxon rank of bahiae) are secondary concerns compared to maintenance of broad mexicanus.”
Comments
from Robbins:
“Clearly Part 1 is straightforward, thus a "YES",
for splitting Sclerurus mexicanus
into at least two species, S. mexicanus
and S. obscurior.
Part IIA. Given this is outside of our region, I presume we are
not voting on this. Nonetheless, genetic data clearly show there is a deep
split between mexicanus and pullus (as compared to other taxa in
this group), so despite similarity in plumage and vocalizations pullus should be recognized at species
level.
I believe d' Horta et al. (2013) and Cooper and Cuervo have
provided good rationale for recognizing obscurior,
andinus, and macconnelli as species, so a " YES " to Part IIB.
Following Cooper and Cuervo's recommendation, at least for now, I
vote "NO" for part III.
Comments
from Stotz:
“I. YES. This seems
straightforward to me with several datasets showing corresponding separation
between these taxa.
“IIA. I am inclined to
say YES on this split, but it is not a case we actually have to consider given
the currently known distribution of the taxa.
However, the North American committee should probably consider this
issue, especially given that it appears SACC will at very least split obscurior
from mexicanus.
“IIB.
YES.
“III.
NO I can see no reason not to follow the recommendation of the
authors of the proposal based on our current knowledge of the situation.