Proposal (818) to South American
Classification Committee
Split Pyrocephalus rubinus into multiple species
Background:
The
Vermilion Flycatcher is a widespread, common species that forms a monotypic
genus. It is not a species that has stood out taxonomically, other than it
often gets called out as unusual for a tyrannid because the male is so brightly
and distinctively colored. What has also caught the attention of some is that
while male plumage of various geographical forms is similar, the plumages of
females are not, with some being quite distinctive … specifically, those from
the Galapagos islands.
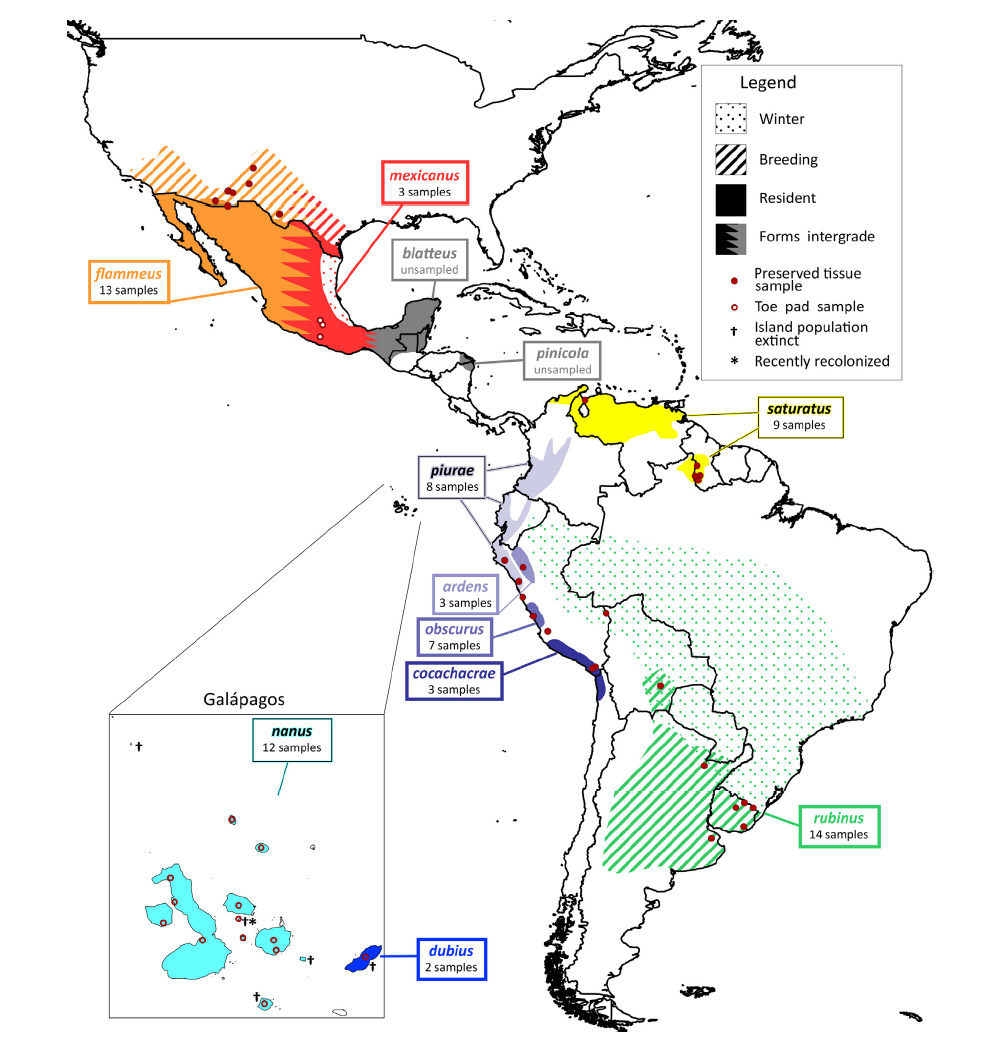
Pyrocephalus rubinus
is very widespread and it shows substantial geographic variation with 12
traditionally recognized subspecies (see distribution map below), much of it
based on differences in female plumage. Although no suggestions to separate the
species into multiples has been made in the past, it is worthwhile to note that
a largely ignored paper by DeBenedictis (1966) notes the radically different
voice of one of the Galapagos populations. DeBenedictis described the aerial
display and vocalization of one population (Isabela Island) in the Galapagos
and confirmed that it is fundamentally distinct from mainland populations.
Rather than a rising series of notes as on the mainland, the Galapagos
population gives a single repeated note. Based on this paper, one would think that
the single species status of Pyrocephalus rubinus would have been called into
question, but as mentioned above this note has largely been ignored, although
recent authors have suggested that P. rubinus is more than one
species (Ellison et al., 2009; Farnsworth and Lebbin, 2004.). Recordings of the
Galapagos birds have not been widely available. In his first trip to Galapagos,
A. Jaramillo was able to obtain poor recordings of the Isabela population of
the Vermilion Flycatcher and confirmed the description of the vocal display as
noted by DeBenedictis (1966).
New Information:
Carmi et
al. (2016) took a fresh look at Pyrocephalus, with molecular datasets in order to
clarify the relationships of taxa within the genus. A total of 85 individuals
was sampled, from 10 of the 12 named subspecies in Dickinson and Christidis
(2014). Two mitochondrial protein-coding genes (ND2 and Cyt b)
were extracted as well as two nuclear loci (ODC and FGB5).
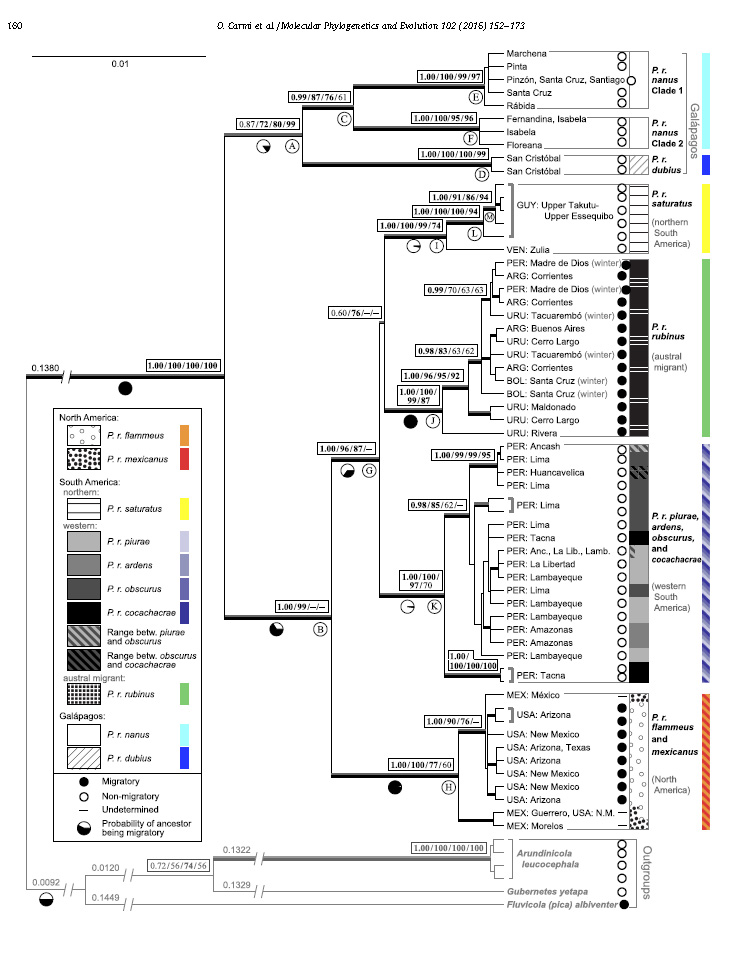
The mitochondrial DNA tree shows that Pyrocephalus
is monophyletic and is separated by a very deep branch from closest
relatives. Seven clades show up in the
data, including three from the Galapagos Islands. These clades from two sister
groups, one of the three clades from the Galapagos, and the other of the
remaining four clades from the continent. In the Galapagos, one clade
corresponds to the subspecies dubius from San Cristobal Island, the geologically
oldest island in the archipelago with a member of Pyrocephalus. The other two
correspond to nanus,
one clade from the older northern islands (Pinta, Marchena, Santiago, Rábida,
Pinzón, and Santa Cruz) and the other from the younger southern and western
islands (Fernandina, Isabela, and Floreana). The continental clades separate
into two groups: a South American group and a North American group. The South
American clade further separates into the austral migratory rubinus
group, the populations along Western South America, and those in northern South
America.
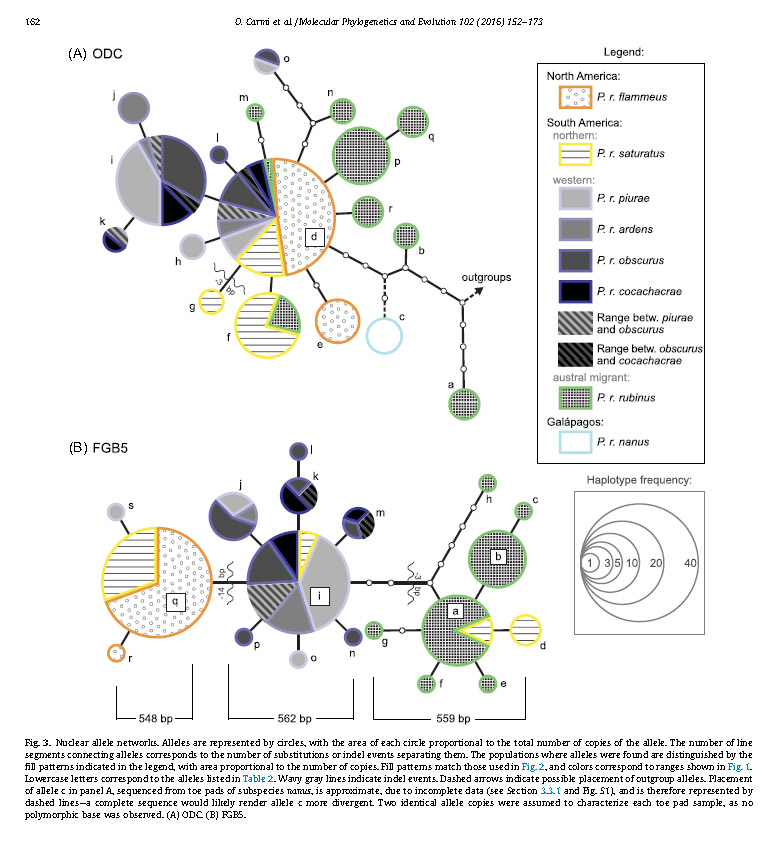
The nuclear allele
networks show a different pattern. In the ODC network, one allele (d) was found
throughout continental populations. Of the five alleles that differed from
allele d by more than one substitution, three were found in Austral rubinus.
The possible root of the allele network was closer to two of these rubinus
alleles. A total of seven alleles were unique to rubinus. One allele was
unique to nanus
from the Galapagos. The highest allele diversity was found in rubinus.
In the FGB5 network there were three groups of alleles, one of several alleles
found in the North American clade, one of birds from the Western South American
clade, and one of rubinus
the Austral migratory clade. Structure
was evident in this dataset; alleles from rubinus were all
characterized by a 3-bp deletion, North American birds by a 14-bp deletion.
Again, rubinus
showed the highest allele diversity of any group. No nanus data was available for this gene.
Galapagos
– the separation between the Galapagos group and the mainland group is
estimated to be roughly a million years ago. These birds are smaller than
mainland birds, with visibly different female coloration. Males are different
as well, with a more restricted red cap than mainland birds. Structurally the
Galapagos birds have very short, weak tails, and short, rounded wings. As noted
above, nanus
is vocally quite different from mainland birds. Their preference for open
forest and forest edge is a habitat quite different from the more open country
taken by the continental populations. Osteological differences have also been
noted and used to suggest species status for Galapagos birds (Steadman 1986).
In summary, various independent lines of evidence can be used to conclude that
there is a different species on Galapagos than the mainland. What is novel is
that the genetic data also clarifies the distinctness of the San Cristobal
population dubius.
Unfortunately, no vocal data is available for dubius, and it may in fact be
extinct now. The branch length separating dubius from the other
Galapagos populations is quite old, suggesting the split is older than any
division seen in the mainland clades. The geographic pattern also fits a
general one seen in the Galapagos, with the old branch (dubius) restricted to the
older eastern islands, in this case San Cristobal. More work is needed to
understand if more than one species is present in nanus, but certainly dubius
appears to be a good species.
Austral migrant rubinus
– There are multiple clades within the mainland Vermilion Flycatchers. Perhaps
there are multiple species level questions to be resolved although nothing
obvious. However, multiple lines of data clarify that the southern migratory rubinus
deserves species status. What is confusing is that the mtDNA data suggest that
it is nested within the mainland group. The nuclear data show a different
pattern where the distinctness of rubinus is perhaps clearer.
There are various reasons why the mtDNA results may be incorrectly showing the
relationship of rubinus,
and on this Carmi et al. (2016) do not
elaborate. The mtDNA data show a sister relationship with the
northern South American group, where rubinus winters. It is not
impossible that historical hybridization with that population may be reflected
in the current mtDNA results and that this may not be its true history? There
is no current evidence for hybridization, and the breeding ranges of rubinus and saturatus do not come close.
The
important point is that the mtDNA do show rubinus to be a separate
clade within the mainland populations. Nuclear DNA further supports
distinctness of this group. But more importantly, the birds themselves show a
clear biological difference, vocalizations. This section was taken out of the
Carmi et al. (2016) paper by the editors. Although sample sizes were low,
playback experiments I have conducted are clear: rubinus does not respond to a
northern song and vice versa. I have more data currently, all of it
unpublished, and the same pattern remains. Furthermore, experiments playing
voice of mexicanus to cocachacrae invoke a response, whereas rubinus is ignored by cocachacrae. The deleted text in the
paper is the following:
“Males
from Belize (subspecies blatteus) were more likely to respond to song from
Arizona males (subspecies flammeus) than to song from Uruguay males (subspecies rubinus;
Wilcoxon signed-rank test W=0, p<0.5, n=10). Males from Uruguay were more
likely to respond to song from Uruguay males than to song from Arizona males
[Wilcoxon signed-rank test W=0, p<0.5, nr=6 (n=9)]. No male from Belize responded to songs from
Uruguay, and similarly no Uruguayan male responded to songs from Arizona .”
The
general nature and pattern of the song is similar in all mainland Vermilion
Flycatchers: a short, rising, and terminally accented trill. The North
American, coastal South American, and northern South American birds have
similar songs. Compared to rubinus they are lower pitched, are delivered more
slowly, and the terminal note is clearly lower pitched than the pitch at the
crescendo of the trill. Here is a typical example: =
https://www.xeno-canto.org/299099
On the other hand, rubinus is higher pitched,
rises quickly and the final note is high pitched, similar to the frequency of
the end of the crescendo. This gives the voice an upwardly accented nature,
quite different from other mainland Vermilion Flycatchers. All can perform bill
snaps during the vocal displays. I have not looked at differences in the call
notes. As noted above, birds of the different song types (rubinus and non-rubinus)
ignore each other’s voice. Given this clear biological response in a suboscine,
rubinus
acts like a good species.
Recommendation:
Based on molecular data, as well as biological (voice)
data, we suggest dividing up the Vermilion Flycatcher into four species: Pyrocephalus rubinus,
Pyrocephalus obscurus, Pyrocephalus nanus, and Pyrocephalus dubius.
Note that rubinus, nanus and dubius would be monotypic. However, P. obscurus would include: obscurus,
piurae, ardens, cocachacrae, saturatus, mexicanus, blatteus, flammeus and pinicola.
English names:
This is
a tough issue as the Vermilion Flycatcher is one of the most widespread and
best known of the Tyrannidae. Although it may be troubling for some to retain
this name for obscurus,
for reasons that have been discussed by this committee elsewhere, in my opinion
the argument for keeping the name is persuasive. Essentially every English
speaker who watches birds in the Americas knows the Vermilion Flycatcher,
changing this name to something else like Northern Vermilion-Flycatcher is
adding complexity to an issue that in the end will create very little confusion
for most people in English-speaking countries. It really is a non-issue for 99%
of the user group of English Names to keep Vermilion Flycatcher even though it
now refers to a subset of what that name used to mean.
I am not
keen on adding a modifier to Vermilion Flycatcher for the various forms and
prefer distinct and evocative names. The easiest of which is to call the
possibly extinct Pyrocephalus
dubius the San Cristobal Flycatcher.
For Pyrocephalus nanus,
the name Galapagos Flycatcher is already taken. In the Galapagos this species
is well known, although it is declining at a precipitous rate. It has become a
conservation concern, and I think to respect what the locals call it, an
evocative name would be Brujo Flycatcher. Locally it is invariably called
“pajaro brujo,” the witch bird. As so many tyrannids have such forgettable
names, why not call the most colorful passerine of the Galapagos by a colorful
name?
Finally, Pyrocephalus rubinus can be
given many names. Perhaps coming up with one that highlights its migratory
tendency, being the only firm migrant within Pyrocephalus is appealing.
But I could not think of any good name that works. I have seen the name
“Scarlet Flycatcher” being used, such as on Xeno-canto. I don’t know if this is
a name they just pulled out of their cloaca or if it has some historical
context? In any case, my preference would be Ruby Flycatcher to match with the
scientific name. Male rubinus are darker than obscurus, a darker red
below and darker brown above. But I don’t think that the color differences are
enough that one could make an argument of ruby or scarlet versus what
vermillion means, essentially, they all suggest a red coloration.
Literature:
Carmi,
O., Witt, C.C., Jaramillo, A. & J. P. Dumbacher. 2016. Phylogeography of
the Vermilion Flycatcher species complex: Multiple speciation events, shifts in
migratory behavior, and an apparent extinction of a Galápagos-endemic bird
species. Molecular Phylogenetics and Evolution 102: 152–173.
DeBenedictis,
P., 1966. The flight song display of two taxa of Vermilion Flycatcher, genus Pyrocephalus.
Condor 68, 306–307. http://dx.doi.org/10.2307/1365570.
Dickinson
and Christidis (2014). 2014. The Howard and Moore Complete Checklist of the
Birds of the World, fourth ed., vol. 2. Aves Press, Eastbourne, UK.
Ellison,
K., Wolf, B.O., Jones, S.L., 2009. Vermilion Flycatcher (Pyrocephalus rubinus). In:
Poole, A. (Ed.), The Birds of North America Online. Cornell Lab of Ornithology,
Ithaca.
Farnsworth,
A., Lebbin, D.J., 2004. Vermilion Flycatcher. In: del Hoyo, J., Elliott, A.,
Christie, D. (Eds.), Handbook of the Birds of the World, Cotingas to Pipits and
Wagtails, vol. 9. Lynx Edicions, Barcelona, pp. 374–375.
Alvaro
Jaramillo, April 2019
Note from Remsen: Voting
structure is as follows:
818A.
Split Galapagos nanus (including dubius) from widespread mainland taxa.
818B.
Treat dubius as a separate species
from nanus.
818C.
Treat all mainland taxa as P. obscurus,
as a separate species from nominate rubinus.
English names (if splits adopted):
818D:
Use separate English names for each species rather than compound names, i.e.
“Something Vermilion-Flycatcher.”
818A-E.
Use “Brujo” as the “first name” for nanus
818B-E.
Use “San Cristobal” as the “first name” for dubius
818C-E1.
Retain “Vermilion” as the “first name” for widespread obscurus
818C-E2. Use “Ruby” as the “first name” for nominate rubinus
Comments from Areta:
“818A. YES to recognizing both Galapagos forms as a
separate species from mainland taxa. The few vocalizations of nanus
I´ve heard are clearly different from rubinus and obscurus,
and the females differ notably from those elsewhere.
“818B. NO, until biological data such as vocalizations (if
not extinct) or more thorough genetic work provides deeper information on the
genetic architecture of dubius. The age of the split between nanus
and dubius
is not impressive, and given that we do not know what this would mean in terms
of reproductive compatibilities, I prefer to recognize dubius as a subspecies of nanus.
Also, note that the difference between the two nanus groups is quite deep in
comparison to that between rubinus and obscurus, yet we are not
discussing their treatment as different species based only on genetic distance.
“818C. YES. I was skeptical of this given the paraphyly of obscurus
with respect to rubinus
(this is something that I would have like explained in the paper itself).
However, after checking all available recordings of songs of the different
taxa, I agree with Alvaro in that the vocal differences between rubinus
and obscurus
(including from mexicanus
to cocachacrae)
are constant. The lack of answer between blatteus and rubinus,
while mexicanus
responds to cocachacrae
seal the deal for me. There is ample room here to publish these playback
experiments together with a thorough vocal analysis of these taxa. It is
regrettable that the comments on vocalizations and playbacks were taken out of
the ms.”
Comments from Claramunt:
“818A: YES. Very complicated case. I think it is fair to tentatively separate
the Galapagos forms as a different species given their plumage, morphological,
and song differences, and the fact that they form a separate mitochondrial
lineage. So, YES to A.
“818B. NO.
Elevating dubius to species mainly because of high levels of
mtDNA "divergence" is not justified, in my opinion. Despite widespread belief, haplotypes with 2%
“divergence” can perfectly coexist within a single species (see Benham &
Cheviron 2019 Molecular Ecology 28:1765–1783). I would like to see more
evidence regarding this potential split. So, NO to B.
“818C. NO. P.
r. rubinus is somewhat distinctive in song and in the fact that it is an
austral migrant but male plumage is barely differentiated and it is not a
different lineage genetically: its mtDNA is part of the south American
continental genealogy, and it shares nuclear alleles, particularly with P.
r. saturatus. Regarding reproductive
isolation, I don’t think that a female of a different subspecies will ignore a
male of rubinus just because he sounds a little different. She may not
react to the song in isolation, but visual cues seem important in this species.
Therefore, NO to C.”
Comments from Stiles: “The genetic data seem quite clear
in mandating splitting up Pyrocephalus
rubinus into SIX species. To begin
with, the name “Vermilion Flycatcher" is solidly entrenched and applicable
to the complex as a whole, as a hyphenated group name - trying to find separate
names that do not include Vermilion seems a bit silly, as any birder anywhere
will recognize a Vermilion Flycatcher! Henceforth,
I will go through the phylogeny as it stands, suggesting E-names en route. 1)
the oldest split is between the Galápagos group and the continental group, so
at the least, one species must be split for the Galápagos, and the internal
split between nanus and dubius is about the same age as the
oldest continental splits, so clearly two species are justified here. Because dubius
apparently is extinct, San Cristobal V-F is appropriate. I see no great problem
with staying with Galápagos V-F for nanus,
as it is now the only extant V-F there but if one must find another name for nanus, Least V-F at least goes with the
Latin name. 2) The next oldest split is between the North-Middle American and
the South American groups, hence at least both must be recognized as separate
species. The former group could be
called Northern V-F (I think mexicanus
has priority as the Latin epithet). 3) The South American group splits into
three well-defined clades of virtually identical ages. If considered as only one species, Southern
V-F would do for all, but if one splits rubinus
from the others, then three species is the only way to go. 4) Very slightly
older is the split between saturatus and the other two.. genetic data are from
Guyana and W Venezuela, but saturatus
also occurs in NE Colombia, so that its distribution is centered on Venezuela,
hence Venezuelan V-F at least does no violence to its distribution relative to
the other two! Carib V-F could be an alternative. 5) From northwestern Colombia
to SW Peru occurs obscurus (originally named for the localized
melanistic form from C Peru, so forget "Dark V-F" as a useful
option!) Hellmayr's name of Pacific V-F fits pretty well. 6) Finally, nominate rubinus, as the southernmost taxon and
an austral migrant, could be called Austral V-F. As a final comment, I can see
no sense in including the northern group under obscurus, producing a flagrant paraphyly. The song of the
southernmost member of the obscurus
group, cacachorea, may show some
resemblance to that of the northern group, but must have been derived
independently; I regard any resemblance as coincidental, or perhaps the
resurfacing of an ancestral character (seems less likely).”
Comments
from Robbins:
“At a minimum, at least two species should be recognized, Galapagos and
mainland based on the differences in vocalizations and genetics. Genetic data support a mainland split into at
least two species, North and South America.
Based on genetic data and the time axis, if one recognizes North and
South America as different species, then one should also recognize Galapagos dubius as a species. Given the three
options we have been presented, for now, I vote as follows:
“818A. YES.
“818B. YES, based on
comments above.
“818C. NO, for
now. I do support recognizing
North/Central American birds from South America, but recognize that is beyond
the scope of our committee. However, depending on member’s viewpoint, that
element may be important for being consistent on how SACC members vote on the
proposals at hand.”
Comments
from Pacheco:
“The multiple data
available are compelling to separate the Galapagos taxa from those on the
Continent. However, for the reasons listed by Nacho and Santiago, I also prefer
to maintain nanus associated subspecifically with dubius. Agreeing with Nacho, I am particularly
impressed by the constant vocal distinctions between the northern taxa and that
southern (nominate) migratory taxon in the continental bloc. Therefore, my votes are: 818A – YES; 818B –
NO; 818C – YES.”
Comments from Remsen: “A. YES.
All lines of evidence point to species rank, as summarized in the
proposal.”
Comments from Bonaccorso:
“818A. YES. All available evidence (molecular,
song, plumage, osteology) point towards a distinct species.
“818B. NO. Agree with Santiago. Genetic distance
and structure should not be the only criteria for species status.
“818C. Abstain. Neither tree topology nor
nuclear networks show a different enough clade. Also, based on tree topology
only, it will seem odd to call P. rubinus a species and lump all other
subspecies into P. obscurus (then P. obscurus will be
paraphyletic). On the other hand,
migratory species are different in the way they speciate (they may cause
paraphyly on the tree). So, it would be
important to publish those song and playback records and do proper analyses, in
order to make a better-informed decision.”
Comments
from Stotz:
“A. “YES to splitting
Galapagos populations from mainland populations. They separate out genetically, vocally and morphologically.
“B. NO to splitting Pyrocephalus
dubius. This taxon is part of a clade with the rest of the Galapagos
birds. Without some sort of data to
suggest species status, I think this is best treated as a part of a Galapagos
endemic species.
“C. NO to splitting obscurus
from rubinus. The vocal
information is very suggestive, but is not published and conflicts with the
published genetic information. I think I
need to see either more information or a clearer statement of the argument
accounting for the distributional and genetic inconsistencies.
“D. “NO. I think I am with Gary on this. While I generally prefer not to create new
compound names, the fact that Vermilion Flycatcher is a very distinctive
flycatcher with a distinctive and good descriptive name, I would like to hang
on to Vermilion Flycatcher for the whole group.
Brujo Flycatcher seems like a mistake.
There is no geographic or descriptive information in it, and Brujo is not
an English word. Ruby Flycatcher is not
too bad, but feels like we are forcing it.
One problem with using Galapagos Vermilion-Flycatcher, given the way the
voting is going, is that then we need a name for the entire mainland
group. Only thoughts I have for that are
Mainland Vermilion-Flycatcher or Common Vermilion-Flycatcher, neither of which
are great names (although HBW uses Common Vermilion-Flycatcher). My guess is that this is a short-term problem
because I expect that we will eventually have sufficient data to split up the
mainland forms into multiple species.”
Comments from Zimmer:
“A. YES. I agree with Mark that
this is the minimum that we should do, based upon concordant vocal,
morphological and genetic data that appear unambiguous in supporting a split of
all Galapagos populations from all mainland populations.
“B. NO. Without more data, I’m
inclined to treat this taxon as a subspecies and part of a single clade of
Galapagos birds specifically distinct from mainland birds, particularly given
that there is geographic structure to the genetic data even within nana populations.
“C. YES. I’m more than a little
confused as how to proceed on this, given the limitations of how 818C is
worded. I agree with those who see a
clear North/Middle American versus South American split, so I don’t feel that
lumping all mainland forms into a paraphyletic obscurus, separate from nominate rubinus is the way to go.
And I definitely don’t support keeping all mainland populations (from
North America to South America) together in rubinus. In looking at the clades supported by the
data, and with biogeographical considerations in mind, my gut tells me that
Gary’s approach is probably the correct one with respect to mainland
populations: 1) North & Middle
American birds (flammeus, blatteus, mexicanus, pinicola) as one species; 2)
Pacific Coast of South America populations (piurae,
ardens, obscurus, cocachacrae) as another; 3)
Northern South America populations (saturatus)
as a third; and 4) migratory Austral populations (rubinus) as a 4th, with Galapagos populations (nanus & dubius) representing a 5th
species. This approach does not fit
within 818C as currently constructed, unless one views the separation of a
paraphyletic obscurus from rubinus, as a necessary first step to
further splitting. With that in mind,
I’ll vote YES, based on the vocal distinctions of rubinus from everything else
(including the results of reciprocal playback trials, which, unfortunately,
were edited out of the paper), just to get the ball rolling, and to pave the
way toward further splitting. I agree
with both Gary and Doug that using “Vermilion-Flycatcher” as an English group
name (to be paired with a species-specific modifier) is the way to go, in what
is a demonstrably monophyletic group.
But that would be putting the cart before the horse, since we don’t yet
know which way the committee will go with respect to the number of splits. If I am understanding Van’s instructions
correctly, we will vote on English names once/if the splits are adopted, but
not until then.”
Additional comments from Stiles: “Taking a closer look at
the topology of the genetic results, I note that the branch separating
saturatus from rubinus + obscurus is extremely short and not all
that well supported, such that the relationships of these three taxa almost constitute
a polytomy (calling into question whether the supposed polyphyly of obscurus
if including saturatus really exists). Hence, I think that the most reasonable course
for now is to include all three under rubinus, and await more conclusive
evidence for their relationships. It therefore means that "Southern
V-F" will do for the E-name, at least for now.”