Proposal (989) to South
American Classification Committee
Treat Ocreatus underwoodii
as multiple species
Background: In an effort to align
world lists, WGAC has split the Ocreatus racket-tails into three
species, a move followed by Clements’/eBird and IOC lists. The main reasons for
this move are based on information provided by Schuchmann et al. (2016);
plumages, morphometrics, display behavior, and biogeography. SACC has not yet
addressed the issue and footnote 62a in Trochilidae says the following:
The
two southern subspecies, annae and addae, were formerly (e.g., Cory 1918) each considered a separate species from Ocreatus
underwoodii, but they were all treated as conspecific
by Peters (1945). Ridgely & Greenfield (2001) suggested that addae
might deserve recognition as a separate species. The subspecies peruana
was also formerly (e.g., Cory 1918) considered a
separate species from O. underwoodii,
but they were treated as conspecific by Peters (1945); Cory (1918) used "cissiurus"
for the species name, but not only does peruana have priority but Peters (1945) considered "cissiurus (= cissiura)"
a synonym of peruana. Schuchmann
et al. (2016) provided evidence from plumage and behavior that Ocreatus
underwoodii should be treated as
four species, with the subspecies addae, annae, and peruana
elevated to species rank. SACC proposal needed.
According
to Schuchmann et al. (2016), Ocreatus comprises between two and four
Biological Species. The plumage characters that they deemed important were: 1)
male ventral coloration, 2) female ventral coloration, 3) tibial tuft (“boot”)
coloration, 4) undertail coloration, 5) male tail morphology (rectrix length
and whether rectrices cross or not), and 6) male tail racket (“flag”)
coloration. Morphological characters of interest were: 1) bill length of
white-booted forms vs. buff-booted forms, 2) wing length of white-booted forms
vs. buff-booted forms, 3) rectrix length (and shape) of white-booted vs. melanantherus/peruanus vs.
annae vs.
addae, and 4) flag size of white-booted forms vs. peruanus vs.
addae/annae. Schuchmann et al (2016) provided
observations on the courtship display from four taxa: incommodus (of the white-booted group, N=17), peruanus (of the buff-booted group, N=11), annae (of the buff-booted group, N=6), and addae (of the buff-booted group, N>15), in
which incommodus had a three-stage display vs. a
two-stage display exhibited by peruanus, and a one- or two-stage display
exhibited by annae/addae. According to Schuchmann et al (2016,
these displays “broadly
correspond with the morphological groups, suggesting the existence of at least
three different groups (underwoodii/melanantherus, peruanus, annae/addae).” Finally, Schuchmann et al. (2016)
mentioned that a case could be made for sympatric
occurrence in eastern (Pogio, near Loja) Ecuador for melanantherus and
peruanus
based on specimen material, noting that the specimens of melanantherus
were of immatures, suggesting local breeding rather than vagrancy (which struck
me as an odd statement, as most hummingbird vagrancy seems to be done by
immatures!), but Zimmer (1951) dismissed this case suggesting instead that the
two immature melanantherus specimens were mislabeled.
Using
the Tobias scoring system, Schuchmann et al (2016) concluded that Ocreatus
contained four biological species: white-booted O. underwoodii, and the
buff-booted peruanus, annae, and addae. Schuchmann et al.
(2016) did not delve into vocal characters given that few recordings were
available on Xeno-canto or Macaulay at the time, and it wasn’t clear if the
vocalizations present were comparable (i.e., homologous).
New
Information:
A quick visit to our LSU collection confirms some of the issues outlined above,
and throws a monkey wrench in the works for others. Overall, I see agreement
between our specimens and the comments on ventral/boot coloration in both sexes
across the various populations represented. I also see the shortening of the
rectrices from northern to southern taxa as was pointed out in the paper. The
sticking point is that there appears to be a population that was not assessed
by Schuchmann et al (2016) from Cushi, in eastern Huánuco, and north into the
Cordilleras Divisoria and Azul (extending from SW Loreto and adjacent San
Martin south to eastern Huánuco and Ucayali depts). Interestingly, although the
authors acknowledge the LSUMZ collection in their paper, and specifically
mention the Cushi locality in their Appendix 1, they apparently did not review
the specimens therein very closely, as they overlooked this population, all
specimens of which should have been present during a revision of the LSUMZ collection
in preparation for this paper. This population appears to display intermediate
characters between peruanus and annae, including flags
intermediate in size and shape between peruanus and annae and
that are more iridescent blue than annae (more like peruanus),
rectrix length intermediate between peruanus and annae,
non-crossing racket-tipped rectrices (like peruanus; although the LSU
specimens are prepared such that this character cannot be assessed, I am basing
this character state on the following field images from the Cordillera Azul: https://macaulaylibrary.org/asset/205180711, https://macaulaylibrary.org/asset/205179041, https://macaulaylibrary.org/asset/611086271, https://macaulaylibrary.org/asset/603149731, https://macaulaylibrary.org/asset/171092391, but note that at
least occasionally, peruanus CAN cross its rectrices as seen here: https://macaulaylibrary.org/asset/513125041), annae
typically crosses its rectrices (e.g., near topotypical: https://macaulaylibrary.org/asset/242905071), and the outer web of
the longest rectrices are not paler-edged as in peruanus, but dark, more
like annae (a character mentioned by Zimmer 1951). It is unclear to me
if these specimens from Cushi and the Divisoria/Azul represent a named taxon
(possibly annae, type locality of which is Chanchamayo, Pasco), or not. Our holdings of male specimens from central
and southern Peru are rather sparse and do not answer this question adequately.
Interestingly, Schuchmann et al (2016) place the specimen from Cushi under the
name “annae” and one from “Fundo Cinchana”
(which appears to be near the “Abra Divisoria” locality of our LSU specimen) in
“peruanus” in their Appendix 1, suggesting that they didn’t detect that
these specimens were anomalous to their taxonomy, but rather considered them to
represent two different taxa!
Unlike
at the time of the writing of Schuchmann et al. (2016), the holdings of online
sound archives are much better with regard to voices of these taxa, and whereas
there is still little indication of what sounds can be best considered
homologous, I can, by ear, detect patterns that strongly suggest that the
white-booted forms (O. underwoodii by Schuchmann et al.’s designation)
are vocally fairly distinctive compared to buff-booted forms. To my ear, the
white-booted birds have a “springier” chase call (the rapid, descending trill)
that often accelerates over the course of one trill and tends to be
higher-pitched (e.g., https://xeno-canto.org/168323, https://xeno-canto.org/693722), whereas the
buff-booted birds have a lower-pitched chase call (e.g., https://xeno-canto.org/226843, https://xeno-canto.org/257059, https://xeno-canto.org/251277) with a tighter, more
even-pace (but can have variable speeds, probably depending on emotional
state). Flat-trill calls (usually given when perched or foraging unprovoked,
pers. obs.) are higher pitched in white-booted birds (e.g., https://xeno-canto.org/398384, https://xeno-canto.org/96236) and lower pitched in
buff-booted birds (e.g., https://xeno-canto.org/22910, https://macaulaylibrary.org/asset/513125041). However, within the
buff-booted forms, I cannot find any strong signal to suggest that there are
distinctive vocal groups. Unfortunately, the Bolivian addae, which
appears to have some of the strongest divergence in morphological characters
within this group, is unrepresented by recordings at the time of this writing;
however, I have heard it and was able to identify it as Ocreatus in the
field in Bolivia, suggesting it is not strongly differentiated from the
Peruvian buff-booted forms that I know well.
For
your listening pleasure:
White-booted
forms (O. underwoodii, sensu Schuchmann et al. 2016):
https://search.macaulaylibrary.org/catalog?taxonCode=boorat1&mediaType=audio&sort=rating_rank_desc&view=grid
https://xeno-canto.org/species/Ocreatus-underwoodii
“O.
peruanus”*:
https://search.macaulaylibrary.org/catalog?taxonCode=boorat2&mediaType=audio&sort=rating_rank_desc&view=grid
https://xeno-canto.org/species/Ocreatus-peruanus
“O.
addae”*:
https://search.macaulaylibrary.org/catalog?taxonCode=rubrat1&mediaType=audio&sort=rating_rank_desc&view=grid
https://xeno-canto.org/species/Ocreatus-addae
*
The division of recordings among taxa by both archives appears to contain some
error (for example, recordings from the Cordillera Azul are included under “peruanus”
when they seem best considered “annae” or perhaps an undescribed form,
based on the specimens I discussed above).
Recommendation: I think there is
enough evidence put forth by Schuchmann et al. (2016) to divide Ocreatus
into more than one species. However, I am not sure I agree that the complex
warrants four species based on the publication and on the specimens and
recordings I reference here. Between the plumage characters (boot tuft color
first and foremost) and the display details, I believe that we can comfortably
split the white-booted Ocreatus underwoodii (sensu Schuchmann et
al. 2016) from the buff-booted forms (peruanus, annae, addae). The
display behavior does not support division within the latter group, nor does
there seem to be strong vocal signal to suggest finer divisions therein. So it
falls on morphological characters to decide if the buff-booted forms require
further species-level separation or not. Personally, I am not convinced that
they do, particularly given the apparently intermediate population I mention
above from eastern Huánuco and the Cordillera Divisoria/Azul area. Therefore, I
recommend the following:
A. Split Ocreatus into white-booted (O.
underwoodii, including polystictus, discifer, incommodus,
and melanantherus) and buff-booted (O. addae, the oldest name,
including peruanus, and annae) species. I recommend YES.
B. Split buff-booted
forms into two species: northern O. peruanus and southern O. addae (including
annae), as per the current treatment by Clements’/eBird, IOC, and WGAC.
I recommend NO as per my reservations based on the Huánuco specimens discussed
above.
C. Split buff-booted
forms into three species: northern O. peruanus, central O. annae,
and southern O. addae, as per the treatment recommended by Schuchmann et
al. (2016). I recommend NO, for the same reasons as in “2” and the additional
reason that addae is lacking archived voice information at present.
English
names would have to be considered if any of these changes from a monospecific Ocreatus
should be adopted. The names already used by Clements’/eBird and IOC are:
White-booted
Racket-tail (O. underwoodii)
Peruvian Racket-tail (O.
peruanus)
Rufous-booted
Racket-tail (O. addae)
Schuchmann
et al. (2016) suggested Anna’s R-t (O. annae) and Adda’s R-t (O.
addae).
Should
we decide to follow recommendation 1 above and not split the buff-booted forms
any further, we could use “Rufous-booted R-t” as has been suggested by Ridgely
and Greenfield (2001).
References:
García,
N. C., K.L. Schuchmann, and P. F. D. Boesman (2022). White-booted
Racket-tail (Ocreatus underwoodii),
version 1.0. In Birds of the World (B. K. Keeney, Editor). Cornell Lab of
Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.boorat1.01
García, N. C., K.L. Schuchmann, and P. F. D. Boesman
(2022). Peruvian Racket-tail (Ocreatus peruanus),
version 1.0. In Birds of the World (B. K. Keeney, Editor). Cornell Lab of
Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.boorat2.01
García, N. C., K.L. Schuchmann, and P. F. D. Boesman
(2022). Rufous-booted Racket-tail (Ocreatus addae),
version 1.0. In Birds of the World (B. K. Keeney, Editor). Cornell Lab of
Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.rubrat1.01
Gill,
F., D. Donsker & P. Rasmussen (Eds). 2024. IOC World Bird List (v14.1). doi :
10.14344/IOC.ML.14.1. (accessed 5 January 2024)
Ridgely, R. S., and P.
J. Greenfield (2001). The Birds of Ecuador: status, distribution, and taxonomy.
Cornell University Press, Ithaca, NY, USA.
Schuchmann, K. L.,
Weller, A. A., and D. Jürgens (2016). Biogeography and taxonomy of racket-tail
hummingbirds (Aves: Trochilidae: Ocreatus):
Evidence for species delimitation from morphology and display behavior. Zootaxa
4200:83–108. https://doi.org/10.11646/zootaxa.4200.1.3
Zimmer, J. T. (1951)
Studies of Peruvian birds. No. 61. The genera Aglaeactis, Lafresnaya,
Pterophanes, Boissonneaua, Heliangelus, Eriocnemis, Haplophaedia, Ocreatus, and Lesbia. American Museum
Novitates 1540. http://digitallibrary.amnh.org/bitstream/handle/2246/3700//v2/dspace/ingest/pdfSource/nov/N1540.pdf?sequence=1
Dan Lane, January 2024
Fig
1. Plate 1 from Schuchmann et al (2016) showing illustration of the different
taxa within Ocreatus, and demonstrating important morphological characters.
Note particularly boot color, male tail length, flag shape and color, and
whether rectrices cross or not.

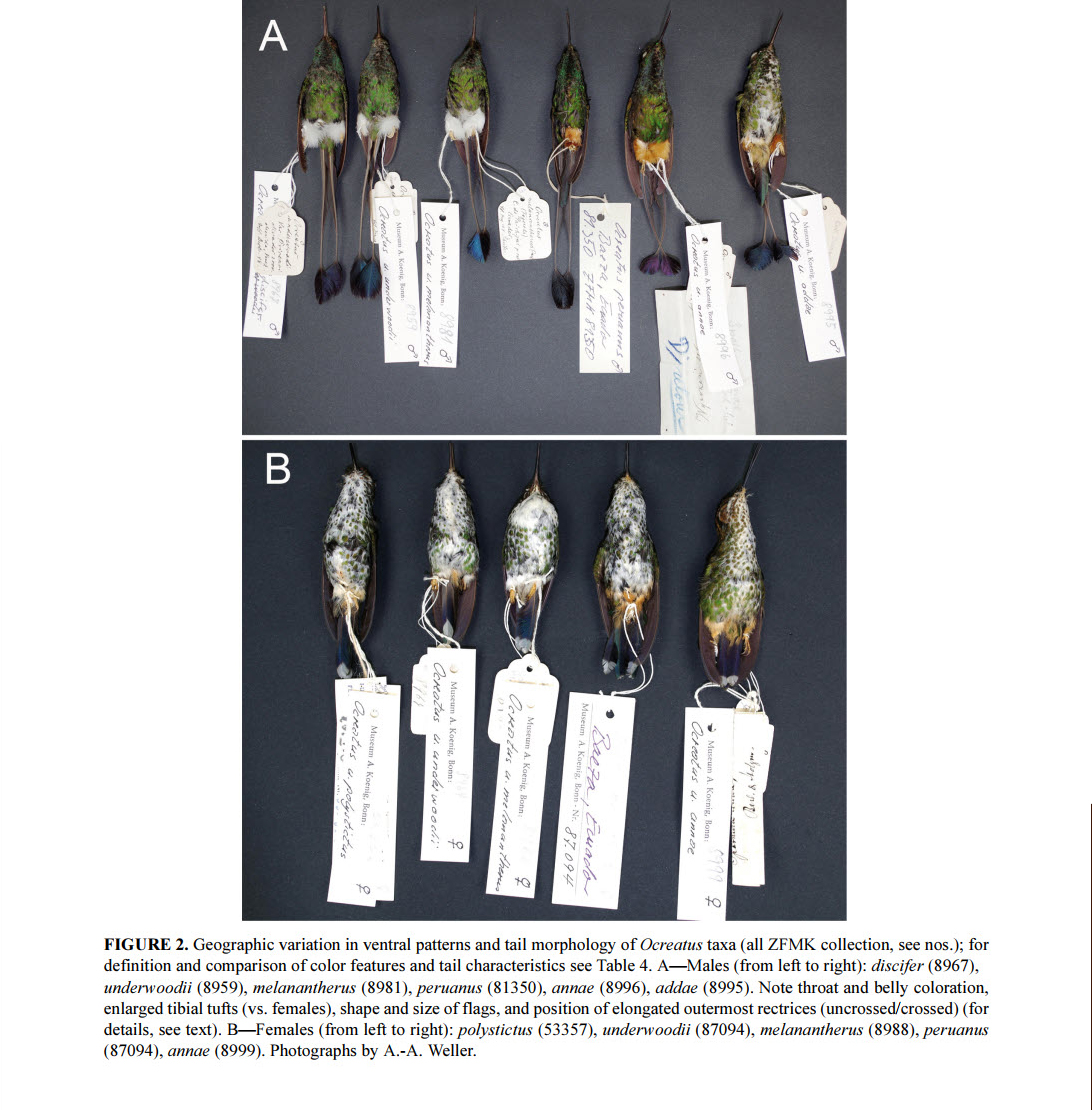
Fig
2. Figure 2 from Schuchmann et al (2016) showing specimens of selected taxa,
including both male and female plumages.
Fig
3: Males of buff-booted forms from Peru and Bolivia, LSUMZ collection. A: peruanus
from San Martin dept (near type locality). B: specimen from Cordillera
Divisoria, Huánuco dept. C: specimen from Cushi, Huánuco dept. D: annae
from Cusco dept. E: addae from La Paz, Bolivia. Photo by D. F. Lane.

Comments
from Zimmer:
“A. YES, for reasons
nicely summarized by Dan. I think the
distinctiveness of the boot-color differences provides an obvious signal,
reinforced by noted display differences.
After listening to archived vocal samples of white-booted birds versus
buff-booted birds, I would agree that the vocal distinctions are also
supportive of splitting these two groups.
“B. NO, based on the
seemingly intermediate specimens cited by Dan in the Proposal, and the lack of
obvious (at least to my ears) vocal differences or display differences.
“C. NO as per my reasons
for voting NO on Part 2, and given that there are no archived vocal samples of
Bolivian addae for comparison with the other buff-booted taxa.
Comments
from Areta:
“In principle, even with notable gaps, the
two-species treatment makes sense, but I will vote NO given several problems,
missing data, and uncertainties.
“The morphological and plumage evidence suggests the existence of
clinality in several features (discussed by Schuchmann et al. 2016). As per the
behavioral data, I am not convinced that the lack of the "whip phase"
(phase C) in the short-tailed forms has been sufficiently corroborated (see
Schuchmann et al. 2016:101 and Schuchmann
1987). For example, intensity and completeness of displays might be
contingent upon the stage of the breeding season, and studying complex
phenotypes for reduced periods of time might not provide thorough descriptions
of displays. Because the last phase apparently leads to copulation, it is
expected that not all the displays will end in this behavior. We don´t have
information on periods of time or places in which these displays were seen.
“The vocal data shows that some calls (whose role in mating is
possible non-existent) seem to differ in pitch. This adds another line of
support on the differentiation between buff-booted and white-booted forms, but
it falls short to show vocal differentiation relevant to mating. Also, note
that the name that takes precedence is addae, but there
are no recordings for this buff-booted distinctive taxon.
“See the relevant part of the tree from McGuire et al. (2014),
which is perhaps the best available genetic evidence, but only includes one
white-booted and one buff-booted taxon, from distributional extremes. The JPLV
sample, which should be JLPV after Juan Luis Parra Vergara [thanks to T.
Schulenberg for working this out], is from Cundinamarca, Colombia, therefore
incommodus? The LSUMZ
sample is from La Paz, Bolivia therefore
addae. The dotted line is at 10
my.”
“This is a case in which more fieldwork, formal bioacoustic
analyses, and genetic analyses are needed to properly assess the taxonomy of
these wonders.”

Comments
from Bonaccorso:
“NO to A, B, and C. I agree with Nacho on all points.
Definitively, genetic data and formal vocal analyses are needed to decide.
Also, a better taxonomic covering regarding displays and probably, more
observations, would further support the case. This is a nice
group of taxa for which a complete phylogeographic analysis is still pending.”
Comments
from Stiles:
“NO for reasons stated above, but additionally for a new wrinkle in the
tapestry – a recently discovered population of buff-puffed birds in the
Serranía de San Lucas, the northernmost spur of the Central Andes of Colombia.
Andrés Cuervo et al. will hopefully obtain genetic data on these (n=3), but
until these are analyzed, it´s impossible to know which (if any) apple carts
might be upset.”
Comments
from Lane:
“I suppose it's no surprise that I'd go with my own
recommendations on this and say YES to 1, and NO to 2 and 3 here. The voice and
plumage distinctions are sufficient for me to feel that a split is warranted.
The news of a new buff-booted form in Colombia doesn't really change that for
me, as it might not be related to the addae group at all. Also, if addae is
distinct from the remainder of the southern buff-booted birds, we can always
act on that once the evidence is provided. Meanwhile, the split seems a
reasonable one to me.”
Comments from Claramunt: “A. YES. At the very least we have two differentiated species here:
white-booted and buff-booted. I am forced to vote NO to B and C as they are
worded, given the existence of intermediate populations revealed by Dan that
suggest that peruanus and annae are not clearly separable. But if given the
chance, I would give species rank to addae, given its distinctiveness in both
male and female plumages, including coloration and tail morphology.
Comments from Niels Krabbe (voting for Robbins): “NO to A,
B and C for the reasons given by Areta. Of all the different characters of the
taxa, only the differences in display strike me as being of more than
subspecific value. Schuchmann et al.'s (2016) sample sizes of observed displays
(17, 11, 6, >15) seem impressive, but do they represent different
individuals, and do they not just reflect levels of excitement?”
Comments from Remsen: “NO to all, echoing especially
the concerns of Nacho and Niels. I
suspect that a two-way split will be justified with more evidence, but I think
a cautious approach at this point is best.
This system deserves a thorough study.
Philosophically, I think this is a good example of why rejected
proposals are in one way more valuable than accepted ones – they not only
provoke additional research to make sure we maximize evidence before making a
taxonomic decision that would potentially signal ”case closed”, but they also
set the stage for additional research by pointing out exactly what we need to
know. This one is a good example – Dan has
laid it all out here in a great proposal that will jump-start further research.”