Proposal (994) to South
American Classification Committee
(Note from Remsen: This proposal was submitted to the North American
Classification Committee and passed, and it is submitted here with permission
from Terry Chesser. This is an
extralimital split, but we have to deal with it because it affects the English
name we use.)
Treat Cattle Egret Bubulcus ibis as two species
Background: Most global lists (e.g., eBird/Clements, IOC,
Howard & Moore) have traditionally considered Bubulcus ibis to be a single species with two subspecies: ibis of southern Europe, Africa, Asia
Minor as far east as Iran, and the Americas; and coromandus of South Asia and southeastern Asia south to Australia
and New Zealand. These two subspecies are separated by a gap in distribution in
Pakistan and Afghanistan. A third subspecies, seychellarum of the western Indian Ocean, is sometimes recognized
(e.g., by Birdlife); otherwise, these populations are considered part of ibis.
The IOC list recently elevated coromandus to species status,
recognizing two species in this complex. Their note on this change is as
follows: “Bubulcus coromandus is
split from B. ibis (Payne &
Risley 1976; McAllan & Bruce 1989; Rasmussen & Anderton 2005). Status
under discussion (Christidis & Boles 2008; Ahmed 2011; HBW).”
The relevant passage from Payne and Risley
(1976), who placed this species in Egretta,
is here:
“Cattle
Egrets of Africa (E. i. ibis) and
India (E. i. coromanda) have very
different breeding plumages and might better be regarded as two species or at
least two allospecies of a superspecies. African birds have orangish-buff
display feathers coloring the entire head, neck, and upper breast; long plumes
of similar color cover the lower back and rump. Indian birds have pinkish-buff
plumes and these are restricted to the crest, the upper breast, and the lower
back; the neck and throat are white. The bill is shorter and stouter in ibis. The extent of feathering on the
tarsus above the distal tarsometatarsal joint is greater in ibis (about 12 mm bare tarsus) than in coromanda (about 24 mm bare tarsus), but
some overlap occurs between specimens of the two groups. Wing lengths differ on
the average (Ali and Kipley, 1968; Mackworth-Praed and Grant, 1970) but the
ranges of wing lengths overlap. The two forms are geographically separated from
each other. Cattle Egrets of the Seychelle Islands have been regarded as intermediate
between the Indian and African birds, but only one specimen in breeding plumage
is known, and it has not been possible to test further the idea that Seychelle
birds (described as a subspecies "seychellarum")
are hybrid results of independent invasions and establishments on the islands
from Africa and India (Benson and Penny, 1971). It is possible that the
differences in breeding plumage would act as behavioral isolating mechanisms
between the two forms of Cattle Egrets, and it would be of interest to
complement the study of behavior of African birds (Blaker, 1969a) with a study
of behavior of birds in India or Australia. Examination of skeletons in the
present study showed no differences in the coded character states in the two
forms, though the interorbital foramen was slightly more rounded anteriorly in
the African specimens.”
McAllan and Bruce (1989), referenced in the IOC
note, is a working list of the birds of New South Wales, Australia. They
presumably recognized B. coromandus as
separate from B. ibis, whereas
Christidis and Boles (2008) presumably treated them as conspecific in their
list of Australian birds.
Volume 1 (Field Guide) of Rasmussen and Anderton
(2005) stated that Western Cattle Egret B.
ibis is “similar but stockier [than B.
coromandus], in breeding plumage with orange-buff mainly on crown, breast,
and mantle.” In Volume 2 (Attributes and Status), they expand on this as
follows:
“[Western Cattle Egret is] Like Eastern but smaller and stockier, with shorter
bill, neck and legs (latter often paler yellowish, olive or grey, but
never black), less bare facial skin and puffier ‘jowls’. Breeding adult
shows a shaggier, paler peach-colored crest only on top of
head, finer, more hair-like peach breast-plumes, and brighter red legs. In
flight, less leg extension than for Eastern….Size Length 330-380 [340-370 in coromandus];
head 90-100 [97-110 in coromandus];
tail 80-90 [81-93 in coromandus];
bare leg 168-180 [205-225 in coromandus]….
Habits Much as for Eastern. Voice Calls noticeably
higher-pitched, more nasal and less gravelly than Eastern’s.?
The Birdlife rationale for continuing to
recognize only a single species was as follows:
“Race
coromandus
The only formal study of phenotypes appears to
be that of Ahmed (2011), a paper published in Dutch Birding and directed
towards identification of potential vagrant coromandus
in the Western Palearctic. He concluded that
“the following features are useful in separating ibis and coromandus: 1 extent
and coloration of adult summer plumage; 2 bill length; 3 tarsus length; 4 tail
length; and 5 bill depth at both nostril and feathering (only in separation
of ‘Indian Ocean specimens’ from ibis and
coromandus). In addition,
vocalisations are of use according to Rasmussen & Anderton (2005) but data
on these were not collected and they require further work. Data to confirm the
validity of the taxon ‘seychellarum’
and its separation from ibis and coromandus are lacking.”
However, although breeding plumage is readily
diagnostic, the morphometric characters listed above, as noted in the Birdlife
spiel as well as by Payne and Risley (1976) and Ahmed (2011), show a fair
amount of overlap (see Table 1 and Fig. 3 from Ahmed 2011 below).
Ahmed (2011) also questioned the vocal
differences discussed in Rasmussen and Anderton (2005), noting that Sangster
(in litt.) could find no differences in vocalizations and that Kushlan and
Hancock (2005) mentioned up to 11 call types. He suggested that the calls
compared by Rasmussen and Anderton (2005) may not have been homologous. I’m not
aware of any further discussion of the vocalizations.

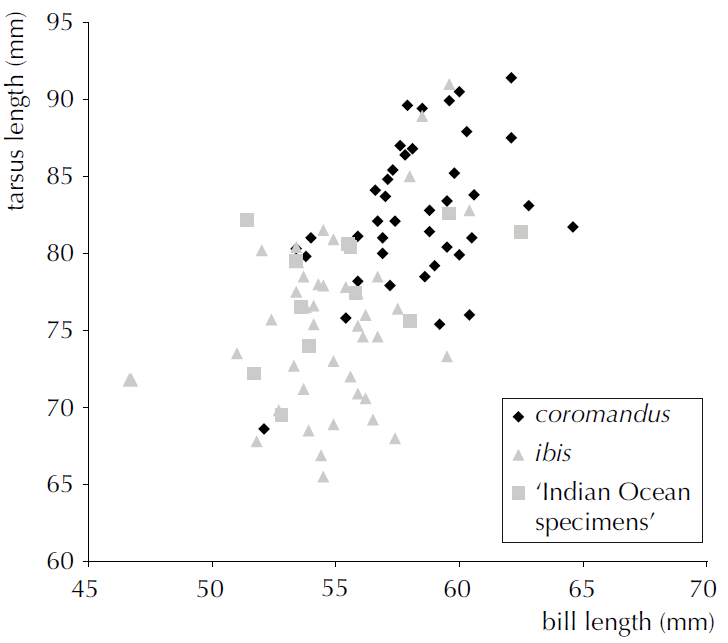
Fig. 3 from Ahmed (2011), plotting bill length
versus tarsus length in the two subspecies of B. ibis, with Indian Ocean populations (“seychellarum”) also separated.
Somewhat surprisingly, no genetic studies have
included both subspecies of B. ibis.
Hruska et al. (2023), for example, included only a sample of subspecies ibis from Louisiana.
New Information:
As part of an effort to consolidate global bird
lists, the IOU’s Working Group on Avian Checklists (WGAC) recently considered
whether to separate B. ibis into two
species. WGAC voted to recognize B.
coromandus as a separate species from B.
ibis. This change has already been adopted in the most recent Clements
update and as noted above, was previously adopted by the IOC list.
Members of WGAC who voted for the split
emphasized the differences in breeding plumage, which involve not only the
extent of the buff coloration but also the color and texture of the plumes.
Also mentioned were differences in shape and proportions, although the
morphometric data do show overlap. The lack of clinality in the plumage
differences was also viewed as significant: breeding plumages of the
westernmost individuals of coromandus and
easternmost individuals of ibis were
noted to be the same as those elsewhere in their respective ranges. Those
voting against the split were not convinced that the differences between coromandus and ibis are more than subspecies-level distinctions, and preferred to
wait for additional data bearing on species status.
Recommendation:
Although I voted against the split, this is
primarily an Old World issue and I recommend that we adopt the new global
taxonomy for this complex, following our standard policy. Most “Old World”
representatives on the WGAC voted for the two-species arrangement. Despite
evidence that may fall short of our usual standards, I would recommend adopting
the new global taxonomy of recognizing B.
coromandus as a species separate from B.
ibis.
English Names:
Both the IOC and eBird/Clements lists are using
Western Cattle Egret for B. ibis and
Eastern Cattle Egret for B. coromandus.
I would recommend that we also use these names, although our guidelines
indicate that the group name should be Cattle-Egret, to indicate their status
as sister species, rather than Cattle Egret.
References:
Ahmed, R. 2011. Subspecific identification and status of Cattle Egret.
Dutch Birding 33: 294-304.
Christidis, L., and W. Boles. 2008. Systematics and Taxonomy of
Australian Birds. CSIRO Publishing, Collingwood, Victoria, Australia.
Hruska, J. P., J. Holmes, C. Oliveros, S. Shakya, P. Lavretsky, K. G.
McCracken, F. H. Sheldon, and R. G. Moyle. 2023. Ultraconserved elements
resolve the phylogeny and corroborate patterns of molecular rate variation in
herons (Aves: Ardeidae). Ornithology 140:
ukad005 doi.org/10.1093/ornithology/ukad005
Kushlan, J. A., and J. A. Hancock. 2005. The Herons. Oxford University
Press, Oxford, U.K.
McAllan, I. A. W., and M. D. Bruce. 1989. The Birds of New South Wales, a Working List.
Biocon Research Group, Turramurra, NSW, Australia.
Payne, R. B., and C. J. Risley. 1976. Systematics and evolutionary
relationships among the herons (Ardeidae). Miscellaneous Publications of Museum
of Zoology 150: 1–115.
Rasmussen, P. C., and J. C. Anderton 2005. Birds of South Asia: The
Ripley Guide. Smithsonian Institution Press/Lynx Edicions, Washington, DC.
Terry Chesser,
March 2024
Note from Remsen on English names. Unless someone wants to write a proposal on
English names with some better names than Eastern and Western, I suggest that
we just go with those to be consistent with NACC and others.
Comments
from Jaramillo:
“YES - I vote to split. We have an issue with herons
and egrets, in that differences and sometimes striking differences, are not taken into account as strongly in birds like these that are
uniform in plumage. White birds, or we could go to Corvus for the
opposite, or Quiscalus. These groups have multiple seemingly
species-level taxa that are cryptic. I am not sure voice is really all that
important in these colonial egrets and herons, in the way it is in bitterns who
use voice as a primary means of attraction and display. We have the
Intermediate Egret system, the Great Egrets, and even the Great White Heron
situation that include some major differences, in display coloration of soft
parts, ecology etc that suggests some barriers to interbreeding. But I guess
our eye goes to the largely white plumage and we stay there. In this case we
have a big difference with their striking difference in breeding plumage, with
extent of color, strength of color, and as noted some of the structure of the
feathers. I think this is important. Similar to the head coloration in Cathartes
vultures, the birds are using these features to sort themselves out. So, I am
fine with a split on the Cattle Egrets.”
Comments from Del-Rio:
“YES. I think plumage differences in breeding forms are pronounced
enough for the separation. Size differences are not as compelling though.”
Comments from Claramunt: “YES. All the evidence points to
separate species. The difference in breeding plumage is remarkable, no
intermediates are known, and the difference has the potential to influence
species recognition. The overlap in morphometrics is not that relevant; sister
species tend to overlap widely in morphometrics.”
Comments
from Stiles:
“YES. The evidence for the split is borderline, but if the WGAC (especially its
Old World members) approved it, I will go along with this.”
Comments
from Bonaccorso:
“YES (but hesitantly). The breeding plumages are different. If that is any
indication of reproductive isolation, it is a good reason to split them.
However, these are the kinds of differences that we consider “subspecific” in
other birds. If there are no intermediate specimens, that could be because they
do not exist, or because the places where they could meet (Pakistan and
Afghanistan) have been rarely sampled. I am also uncomfortable with a split
with no genetic data, but perhaps the taxonomy of the cattle egrets should not
be a big concern. If somebody finds out later that they are conspecific, we
just lump them back again without any major consequences.”
Comments
from Robbins:
“NO. I find current data not to be compelling for recognizing these two egrets
as separate species. For example, I do
have experience with “Eastern” and in at least some populations it does have
black legs (have photos on my eBird checklists that document that) contra what
R & A (2005) state, “never black”.
Moreover, I also question other morphological statements based on my
experience. It should be noted what is attributed in this proposal to the
Birdlife rationale that also contradicts R & A statements on differences in
morphology. The comments on
vocalizations by Ahmed (2011), contra R & A, should also raise a red
flag. I want to see a genetic data set
from across the range of “Eastern” as compared to “Western” before a taxonomic
change is made.”
Comments from
Areta:
“I vote NO to the split. I
would like to see more compelling evidence. The current differences in plumage
(breeding plumage differences especially, other features seem less reliable)
shown do not allow me to judge whether a two-species taxonomy is an improvement
over a single-species taxonomy with subspecies. There is ample morphological
overlap as well. A better study can do no harm, so I prefer to wait for it in
order to revise this position. This is a contentious issue that needs to be
properly studied.”
Comments from Remsen: “NO. These are all
interesting observations on the differences between the two taxa, but they are
largely anecdotal and disputed (voice, texture of breeding plumes ) and the
measurements overlap. That leaves
breeding coloration. I’m not sure this
marks the “point of no return” in restricting gene flow heron speciation. Certainly, we have color morphs within some
species (Egretta rufescens, E. sacra, E. garzetta) that suggest
that major differences in plumage coloration are not barriers to gene
flow. On the other hand, we have Ardea
herodias/A. occidentalis that mate assortatively at their contact
zone (yet continue to be treated as conspecific by NACC), for which the only
difference is coloration and thus contradict the point made about color morphs. For better or worse, global Butorides
striatus taxa ranked as subspecies differ at least as much in plumage from
each other as do the two Cattle Egret taxa.
Nycticorax caledonicus also.
In my view, species limits in allotaxa in the Ardeidae is an
inconsistent mess with not many “knowns” in terms of contact zones in
parapatric taxa to provide any guidelines.
So, strictly as a matter of principle, I don’t think we should be
changing long-standing species limits without a better empirical and
theoretical framework.”
As
pointed out to me by Murray Bruce, Vaurie (1963, BBOC, “Systematic notes on the
Cattle Egret (Bubulcus ibis)” was one of the few people who really
studied specimens; he did not make a recommendation to treat them as separate
species but provided a good synopsis of the differences:

“Tangentially,
Vaurie makes the point that I’m sure we’ve all pondered. Why is there such a big gap in distribution
between these two taxa, which have expanded dramatically with the spread of
livestock and agriculture, and which are notoriously good at dispersing, most
notably in crossing the Atlantic?
“I
appreciate that good field people familiar with coromandus say that it
flies differently and has a different “look”.
Maybe after just a day of field experience I would be impressed by these
differences. Nonetheless, the actual
data for changing the taxonomy is weak at best.
In a comparative context, I can’t think of another pair of sisters in
herons that differ solely in extent of a single color in the plumage during the
breeding season. I’d be more impressed
if there were differences in breeding facial coloration and bill color, but
even those differences are on both sides of the species/subspecies line in
current taxonomy.”
Comments
from Bonaccorso:
“I change my vote to NO in light of the other comments. My YES was reluctant,
anyway.”
Comments from Zimmer: “NO. I would echo some of the same caveats already
presented by Mark, Nacho, and Van on this one.
The distinctions in breeding plumage between ibis and coromandus
are suggestive, but I would find them much more compelling if there was also
clear-cut evidence of difference in bare parts coloration when in peak breeding
condition. To me, the differences in
facial skin coloration, bill coloration and tarsus & toe coloration are
among the most dramatic morphological changes in this group of birds tied to
the peak of breeding condition. Changes in the colors of these bare parts are
not only dramatic, but are highly seasonal, and highly ephemeral, and are
coincident with “peak breeding condition”, which would seem to make them all
the more likely to be important as isolating mechanisms and in mate
selection. There may, indeed, be
distinctions in bare parts colors between ibis and coromandus,
but, if so, I don’t see any indication of it in the Proposal or in any of the
excerpts from the relevant literature cited therein, and my own field
experience with coromandus is too seasonally limited to say.”
Comments from Lane: “YES, although it’s
mostly because this is an Old World issue, and the result is immaterial to the
SACC region. As others have mentioned, I am not sure what characters are
informative in many heron species limits cases, so if this pair truly is that
distinctive or not is not clear to me. There are simply too few recordings of B.
coromandus to have any useful idea of how similar or different they are
from B. ibis (especially given the dilemma of homology of vocalizations?).
However, I have been thinking that, if the split is to be, we could maintain
“Cattle Egret” for B. ibis, and use “Buffalo Egret” for B. coromandus,
given that is the Bos species they probably evolved alongside (that is
only partly tongue-in-cheek).”