Proposal (1003x) to South
American Classification Committee
Species limits in the Myioborus
melanocephalus complex, revisited.
As
originally presented, this proposal (see below) did not pass, but it was
suggested that I present a revised version with more details on specific parts
of the proposal, progressing from north to south.
A.
Continue
to recognize M. albifrons as a species distinct from the rest of the
taxa related to the hybrid zone. The alternative would be effectively lumping
all of the complex into a single species ornatus, which has priority (a
possible option suggested by two members of SACC). However, no evidence of
hybrids between albifrons and ornatus exists; moreover, recent
eBird records of albifrons from the Tamá area on the border between
Colombia and Venezuela effectively establish parapatry between albifrons
and ornatus which would render untenable lumping them. Hence, I
recommend NO for this option.
B.
Recognize
M. chrysops as a distinct species from M. ornatus. The original
proposal failed to pass principally because of doubts regarding this split.
However, it was handicapped by the rather poor illustration of chrysops and
the lack of a good illustration of M. o. ornatus. Here are two photos
that I think represent these taxa more clearly:
M. o. ornatus:

M. chrysops:

Both taxa show a
generally similar pattern of brightly colored faces that present strong
contrasts with the dark irides, and with black napes, set off from dark gray
backs and wings. In both, note a fine white line in the black of the side of
the neck, approximately coinciding with the lower posterior border of the
auriculars. These patterns occur throughout the respective distributions
of each taxon. The principal difference between them is in coloration of the
face: glossy white in ornatus vs. glossy orange-yellow in chrysops. The
color of the underparts of ornatus is bright yellow, as are the forehead
and crown; chrysops is a more orange-yellow below and has a notably
different distribution of colors on the head, with the orange forehead much
more prominent and extending back over most of the orange-yellow crown. The current distributions of the two do not
overlap: chrysops occurs widely in the Central Andes, wherever the
elevation extends well above the 2000 m ridgeline – essentially an archipelago
of high Andean islands – and more locally in the Western Andes, where such
islands are much fewer and more isolated. Note that the isolated population of chrysops
where the Eastern Andes unite with the Central Andes is isolated by ca. 100
km from the southernmost limit of ornatus and is ca. 50 km east of the
hybrid zone between chrysops and the melanocephalus group. With
the current global warming, the geographic separation of chrysops and ornatus
can only increase, s.s.as the white postauricular line) do not represent
current or recent hybridization but more likely, the retention from a
considerably earlier common ancestor. To summarize:
The hybrid zone in the melanocephalus complex is strictly
with chrysops: ornatus is not directly involved. It thus more
clearly expresses the established precedent that the hybridization is occurring
between two full species. The distributions of ornatus s.s. and chrysops
are separated completely by at least 100 km of unsuitable habitat and no
hybrids between them are known. Their plumages differ strikingly in the head
region, of a magnitude similar to those distinguishing many other species of Myioborus;
plumage similarities between them are not evidence of current or recent
hybridization but retention of more ancestral characters. Perhaps pertinent
here is that the phenotypic differences in color and pattern between ornatus
s.s. and chrysops are if anything greater than the differences
between ornatus s.s. and albifrons, which appear to be parapatric
species.
Points in favor of a NO: The shallow
divergence in mitochondrial genetics between both species, although this could
be invoked for the complex generally, including M. albiceps, which is a
close sister to the remaining members, but which is apparently parapatric with M.
o. ornatus. The lack of direct evidence that the latter could interbreed
with chrysops were they to enter into contact is a piece of the puzzle missing, given their
disjunct distributions of some antiquity and is thus impossible to resolve. A
NO vote here would imply that the entire complex represents a single species
and consequently, the hybridization event occurs between subspecies of a
phenotypically extraordinarily diverse species.
My personal opinion
here favors a YES.
C.
Suppress
the name ruficoronatus due to the hybrid nature of its type, thus
rendering this name inapplicable to any described taxon. This should be an
undisputed YES given its clear genetic identity and distribution.
D.
Select
bairdi as the second parental species involved in the hybridization,
reflecting its adjacent distribution, its stable phenotype through central and southwestern Ecuador and the
compatibility of its phenotype for integration with the hybrid zone. This should
be an easy YES: no other taxon unites all of its qualifications.
E. Recognize griseonuchus
as a separate species from bairdi. At its southern limit around the
Ecuador-Peru boundary, bairdi meets griseonuchus, a poorly known
taxon that is phenotypically most similar to bairdi but the two are
reciprocally diagnosable. The current treatment appears to favor treatment of griseonuchus
as a subspecies of bairdi, but apparently there is no evidence of
hybridization between them, but only further collections and observations can
fully resolve its status as a subspecies or separate species.
I
lean toward a YES here, but tentatively, pending more data from the potential
zone of contact between them.
F.
Split
the remaining southern members of the complex as a separate species, M.
melanocephalus. This reflects the fact that the three included species have
black crowns, and zoogeographically, all occur east of the río Marañón valley
whereas bairdi and griseonuchus occur to the west; this low,
relatively dry valley is well recognized as a major barrier for the
distributions of taxa of the wet highland forests of opposite sides of the
valley. This decision should logically be YES.
G.
Continue
to recognize the currently known subspecies within M. melanocephalus. Its
three subspecies are distributed sequentially from northwest to southeast,
collectively ranging from northern Peru south to central Bolivia: malaris,
melanocephalus and bolivianus. Cuervo and Céspedes presented brief
descriptions, but emphasize that their respective distributional limits are
poorly documented, and the genetic characterizations are relatively incomplete;
clearly this group merits further study, but for the present they considered it
best to continue recognizing all three as described. I recommend a YES here.
Note from Remsen: At Dan Lane’s
suggestion, here are the published trees for Myioborus in case they are
helpful. Both are based on mtDNA
sequence data:
Lovette et al. (2010; MPE):
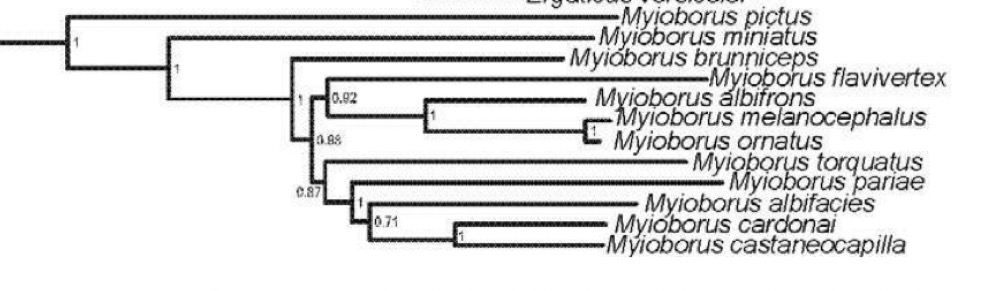
Pérez-Emán (2005; MPE):

Gary Stiles, September 2024
Voting chart: https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Zimmer:
“
“A.
I am a little confused by how this one is stated in the Proposal. It begins, by saying “Continue to recognize M.
albifrons as a species distinct from the rest of the taxa related to the
hybrid zone.” I would vote YES to
that for the reasons stated by Gary in the sentences that follow (e.g. the
alternative would be lumping all of the complex into a single species ornatus…
but there is no evidence of hybrids between albifrons and ornatus…and
eBird records effectively establish parapatry between albifrons and ornatus,
which “would render untenable lumping them”), but then, he says “Hence, I
recommend NO for this option.” It seems
to me that Gary is recommending NO to the option of lumping all taxa in the
complex into a single species, ornatus, to which I would also vote NO,
but the initial question was whether to continue to recognize albifrons
as a distinct species, to which I would reiterate my YES vote. Or am I missing something here?
“B. Recognize M.
chrysops as a distinct species from M. ornatus. I’m persuaded by the clarification presented
in the revised proposal, so a YES for me on this one.
“C.
Suppress the name ruficoronatus due to the hybrid nature of its
type. This would seem to demand an
obvious YES.
“D.
Select bairdi as the second parental species involved in the
hybridization. Another obvious YES.
“E.
Recognize griseonuchus as a separate species from bairdi. I’m uncommitted on this one. I think I would need more information to pull
the trigger on splitting these, so I lean toward a NO for now vote. The same logic that calls for treating the
three black-crowned taxa that occur east of the rio Marañón valley as a single
polytypic species (see Part F), would seem to argue for treating rufous-crowned
bairdi and griseonuchus from west of the Marañón as a single
polytypic species, rather than as two distinct species that are similar but
diagnosable.
“F.
Split the remaining southern members of the complex as a separate species, M.
melanocephalus. YES.
“G.
Continue to recognize the currently known subspecies within M.
melanocephalus. YES.
“What
I don’t see here, is a sub-part of the Proposal that explicitly addresses the
question of bairdi versus chrysops, given the broad hybrid zone,
with these two as the sole recognized parental types. If we are treating them as a single species,
and, if griseonuchus is treated as a subspecies of bairdi, then,
by my count, my votes would support recognition of 4 species: albifrons, ornatus, chrysops (including
bairdi and griseonuchus), and melanocephalus (including 3
sspp), which, I believe, is what both Mark and Dan suggested as an alternative
in the original Proposal. That would be
my leaning too, for 4 species.”
Comments
from Bonaccorso:
“Based on the logic presented in the reappraisals above, recognizing four
species seems reasonable:
“1. Myioborus
albifrons: Phylogenetically distinct (long mitochondrial DNA branch in
Lovette et al, 2010 and Pérez-Emán 2025 papers) and phenotypically diagnosable.
2. Myioborus ornatus:
Not really part of the hybridization issue (since the hybridizing forms are chrysops
and bairdi). Also, phenotypically diagnosable and certainly does not
hybridize with neither albifrons (according to information provided by
Gary) nor chrysops.
3. Myioborus
chrysops: Includes chrysops, bairdi, and griseonuchus until
the hybridization zone is better understood with nuclear data. The geographic
continuity among these forms suggests a fluid entity composed of three
subspecies.
4. Myioborus
melanocephalus: Encompasses would include malaris, melanocephalus,
and bolivianus. According to Céspedes-Arias et al (2021) these three
forms share a fairly connected mitochondrial DNA haplotype network.”
Comments
from Stiles:
“YES to ABCDE - all the splits. Only one suggests very restricted gene flow
(which in relation to the broad expanses in the distributions of each showing
none, suggests menor preocupatión). Should any additional exchanges be
discovered, the zone of hybridization will almost surely be very narrow. An excellent study.”
Comments
from Areta:
“I developed the logic of my arguments before, and I still stand by them. I
find the proposed voting issues and sequence to be convoluted and unnecessarily
complicated. I think that the most
reasonable course of action based on current evidence is to recognize three
species largely following a N-S replacement: M. albifrons, M. ornatus,
and M. melanocephalus. Vocal and
genomic data will surely provide a richer perspective, and while I do not claim
that the 3-species solution is definitive, it is the one I see as currently
more defensible when integrating genetic patterns of differentiation, phenotype
(plumage and vocalizations), and hybrid zones:
A.
Myioborus albifrons: YES,
very distinct and parapatric to ornatus
without known hybrids. I do not agree with Gary´s assessment indicating that
"the phenotypic differences in color and pattern between ornatus s.s. and
chrysops are if anything greater than the differences between ornatus
s.s. and albifrons". Instead, I think that albifrons is very different from ornatus, while ornatus
and chrysops are very similar
(compare the photograph below to those of ornatus
and chrysops that Gary shared).

https://macaulaylibrary.org/photo/88090771
B.
Myioborus chrysops: NO. Include
the chrysops complex (chrysops,
bairdi, and griseonuchus) together with ornatus which has priority.
B.
Addendum. Myioborus ornatus: YES,
including chrysops, bairdi, and griseonuchus given the
geographically widespread and rampant hybridization, and also of course ornatus (given the broad similarity in
plumage and vocalizations to the chrysops
complex as argued previously in my first vote). As mentioned in point 1, ornatus is just slightly different from chrysops (slightly darker yellow, with
reduced white on the face), and under the BSC/Recognition Concept I don´t think
they will keep their integrity: I predict rampant hybridization would happen if
they ever meet, even if their current ranges are now allopatric.
C.
YES. ruficoronatus represents a hybrid phenotype and I don´t see a need
to keep recognizing it.
D.
NO. It is clear that bairdi
is the second taxon involved in the hybrid zone, but given the geographic
breadth and frequency of hybridization, I don´t support recognizing bairdi
as a full species. Instead, I consider it a subspecies of M. ornatus (see B and B addendum) that hybridizes with the
subspecies chrysops.
E.
NO. I don´t see strong evidence to
recognize this taxon as a full species (see F).
F.
Myioborus melanocephalus: YES
(although there is room for debate), including malaris, melanocephalus,
and bolivianus. Note that, as discussed before, the situation across the
Marañón separating griseonuchus and malaris is not clearcut based on genetic
information, and vocally the melanocephalus
group is pretty much like ornatus
(including the chrysops group). I
vote to retain melanocephalus for the
time being, but I would not be averse to lumping it with ornatus if genomic+vocal evidence arise supporting such a lump.
G.
YES, as in F.
Comments
from Jorge Pérez-Emán (voting for Del-Rio): “This proposal to define species
limits in one monophyletic group of the genus Myioborus is full of complexity. The evolutionary history and
differentiation of the northern/central high Andean species complex (albifrons, ornatus and melanocephalus)
is a fascinating system to study evolutionary processes associated with the
generation of diversity. Earlier studies provided a phylogenetic hypothesis for
the group and suggested potential paraphyly for melanocephalus in relation to ornatus
(Pérez-Emán 2005, Lovette et al.
2010). This finding associated with the increasing information of observations
of phenotypically intermediate birds in southern Colombia – northern Ecuador
led to the study of Céspedes-Arias et al.
(2021) that described and analyzed the complex system of hybridization between
these taxa. Cuervo and Céspedes (2023) followed up with a revision of the
taxonomy of the group and provided with different proposals congruent with
findings of this last study. These studies underscore a complex evolutionary
dynamic that generates lots of questions, but we are still short of having
clear answers. Consequently, when I see a proposal (and subsequent
responses/votes) suggesting one (just as the baseline), two, three, four, five
and six species (from a universe of nine voters), it suggests that uncertainty
is larger than the information required to make clear and stable changes to the
current taxonomy of the group. I will share my thoughts (and available data)
for each of the proposal optional splits and then summarize/ponder the
different scenarios for number of species suggested for this group of Myioborus in light of what we know right
now.
“A. Myioborus albifrons, a separate species?
This is a taxon restricted to the Venezuelan Andes (Tachira, Merida and
Trujillo) and it is allopatric in
its distribution in relation to other members of the group (specifically, M. o. ornatus), with no records of
hybridization between these taxa. These taxa are potentially isolated by the Táchira
Depression, and arid valley separating the Venezuelan Andes from the Tama
mountains from where, as far as I know, there are no records of albifrons. Phenotypically it is
characterized by a crown black with a rufous patch with several feathers tipped
black and white forehead, supralorals and eyering, forming a “white spectacle”,
a plumage pattern not very different to that of melanocephalus populations north of the Marañon River (Figure 1).
Molecular studies suggest M. albifrons
is the sister taxon to the ornatus-melanocephalus
complex and diverge from them around 4% uncorrected sequence divergence in some
mtDNA genes, with a small intraspecific genetic variation (Pérez-Emán 2005).
Such phylogenetic relationship was supported by increasing number of both
mitochondrial and nuclear genes (Lovette et
al. 2010) and a recent mitogenomic analysis (Zhang et al. 2025, which
unfortunately sampled ornatus from
toepads, and their nuclear results were messy in this case). In summary, I think
all data currently available support species status for this taxon.
“Figure 1. Plumage color variation in M. albifrons and M. melanocephalus north of the Marañon River. Notice the
similarities in plumage coloration pattern between both species, the intensity
variation in the ventral yellow coloration (in all taxa), and the absence of
black nape and variation in size and color of crown patches throughout the
distribution of melanocephalus.
Top row: M. albifrons (Venezuela) from
left to right: Táchira, https://ebird.org/checklist/S220813847; Mérida,
https://ebird.org/checklist/S162418134; Mérida, https://ebird.org/checklist/S220813847.
Middle row: M. melanocephalus “ruficoronatus” (bairdi, Ecuador), from left to right, Pichincha,
https://ebird.org/checklist/S162780652; Tungurahua,
https://ebird.org/checklist/S138756945; Tungurahua,
https://ebird.org/checklist/S160083246; Azuay, https://ebird.org/checklist/S113062604; Loja, https://ebird.org/checklist/S152346432;
Third row: Myioborus melanocephalus
griseonuchus (Peru), from left to right: Piura,
https://ebird.org/checklist/S124270377; Cajamarca,
https://ebird.org/checklist/S61949069; Cajamarca,
https://ebird.org/checklist/S65191736; Cajamarca,
https://ebird.org/checklist/S60936330;

“B. Myioborus ornatus: two species (ornatus and chrysops)? Previously considered geographical variation of M. ornatus, labeled subspecifically,
these taxa were recently proposed to represent different species by del Hoyo
and Collar (2016), using the Tobias et al.
(2010) phenotypic criteria for species delimitation. They justified such split
based on differences in the white vs. yellow around the eye, amount of yellow
and its intensity on the forehead and ventral side, respectively, and slight
differences in the length of vocalizations. Additionally, this proposal
includes the geographical isolation of o.
ornatus and o. chrysops and the
isolation of o. ornatus from the
hybrid zone between o. chrysops and melanocephalus. It is emphasized that
the hybrid zone is strictly with o.
chrysops and not o. ornatus and
that no hybrids between these taxa are known. It adds that phenotypic
differences between o. ornatus and o. chrysops are about the same or larger
than between o. ornatus and albifrons.
“Although all these points are potential good
reasons to consider each taxon as separate species, it is important to consider
phenotypic variation, mostly plumage here, but also vocalizations. As Nacho
indicated previously, vocalizations are very similar in all this group (even albifrons) and slight differences in
length of the song, shorter or longer than 5s (even considered minor
differences by del Hoyo and Collar 2016) could also be the result of sampling
size and geographical biases (which I cannot confirm because I did not find the
data supporting these differences). On the other hand, pictures of these taxa
taken throughout their distribution (Figure 2) show an amount of plumage color
variation that suggest plumage differences are not clear-cut, even though we
could find good examples of ornatus
and chrysops in the northern portion
of each taxon ranges (but no rigorous data available at this moment). Moreover,
although I could not find on my files the complete specimen information, many
years ago I found at the Instituto de Ciencias Naturales de Bogotá, Colombia
(ICN), one specimen of ornatus
collected by Bernal and Olivares (potentially from the 60’s?), from Fusagasuga,
Cundinamarca, that approaches the plumage pattern of “typical” o. chrysops, side by side with other
individual “typical” o. ornatus from
the same locality (Figure 3). Although these pictures/specimens do not
represent the common pattern throughout the distribution of each taxon, they
show that there is variation in their plumage, that is “possible” to go from
one plumage type to the other, and that individual variation could even be
larger than among locality variation (see Céspedes-Arias et al. 2021).
“Figure 2. Plumage color (and pattern)
variation in Myioborus ornatus from
Colombia. See variation in ventral coloration (bright yellow to orange in both
taxa) and the amount of white in the face of o. ornatus (even almost disappearing in one individual from Bogotá).
Notice that variation is not geographically structured as some of the
differences are found at the same or nearby localities.
Top row: Myioborus ornatus ornatus
from left to right: Bogotá, https://ebird.org/checklist/S38604592; PN Chicaque,
Cundinamarca, https://ebird.org/checklist/S202692769; Bogotá,
https://macaulaylibrary.org/asset/192970091; Bogotá, https://ebird.org/checklist/S103828264.
Bottom row: Myioborus ornatus chrysops,
from left to right: Antioquia, https://ebird.org/checklist/S124051776;
Antioquia, https://ebird.org/checklist/S121272164; Antioquia,
https://ebird.org/checklist/S100185842.

“Figure 3. Pictures of Myioborus ornatus ornatus from Fusagasuga, Cundinamarca, Colombia.
The specimens are from the collection of the Instituto de Ciencias Naturales (ICN), Bogotá,
Colombia (numbers 1425 and 5310). It is far from a good picture, but it shows
within locality variation in face pattern/color of this taxon in the range of o. ornatus. The specimen with less white
(#5310) was collected by Olivares and Bernal.

“We also need to focus on the data shown by
Céspedes-Arias et al. (2021) in
relation to the hybridization pattern between these taxa. Phenotypic variation
is huge and clearly suggests a hybrid zone that includes southern Colombian and
northern Ecuador. However, the hybrid zone was described based on the
phenotypic characters, but the other part of the story is that there were no
fixed haplotypes at either end of the hybrid zone that could be used to
characterize the parental populations. I think information from phenotypic
characters in these hybrid zones is of great importance (as it is also the
potential subject of selection), but it is also important to consider that the
hybrid zone could be different if we focus on the genetic data. Extensive
sampling of mtDNA (ND2 gene) variation shows that the most common haplotype is
found throughout the complete distribution of both o. ornatus and o. chrysops
and reaches Tungurahua, Ecuador, in the range of melanocephalus. Moreover, out of 27 o. ornatus samples, 14 of them shared haplotypes with hybrids.
Similarly, 10 out of 15 samples of o.
chrysops were characterized by the hybrid haplotypes; and we are referring
to “phenotypically pure” diagnosed taxa. We could interpret these findings as a
really young divergence between these taxa (recent gene flow between them), and
that both ornatus taxa are involved
in the hybrid zone with the presence of genotypic hybrids. Lastly, but not less
important, Céspedes-Arias et al.
(2021) indicated the presence of concealed black feathers with rufous base far
from the hybrid zone in both o. ornatus
and o. chrysops.
“In summary, given the clear similarity between
both taxa plumage pattern and coloration, as well as vocalizations, and the
fact that they are genetically homogeneous throughout their distribution,
including the hybrid zone, the evidence supports more a scenario of a single
species with geographical variation rather than separate species. However, how
could such phenotypic differences exist in the presence of such similarity at
the genetic level? more of that below but, for now, my take would be of one species
including both taxa.
“C. Myioborus
melanocephalus ruficoronatus is not a valid taxon. The evidence presented
by Cuervo and Céspedes (2023) is thorough and definitely supports the proposal
of considering ruficoronatus a
non-valid taxon and assign the populations of birds with such similar phenotype
to bairdi. I indicate similar
phenotype because the ruficoronatus
type has similar plumage coloration to hybrids in the region, but not the same
as the population we historically have named ruficoronatus. Consequently, the type is proposed to be a hybrid
and the locality type reassigned to Pasto (no evidence of Cali as collection
site). The history of several of these old types is not always simple but
rather convoluted, and I think the work by Cuervo and Céspedes (2023) is
thorough and complete as it could be with the evidence on hand. I just want to
emphasize, due to potential misinterpretation from the proposal, that there are
no genetic data from the type, and its distribution is inferred based on
coloration patterns from the hybrid zone by Céspedes-Arias et al. (2021).
“D
and E. Select (and recognize) M. m.
bairdi as the species (taxon) hybridizing with ornatus and include griseonuchus
in this species (or recognize it as a different species) I include both
sections of the proposal here as they are really interconnected. If we accept ruficoronatus should be replaced by bairdi as the appropriate name for the
adjacent form to ornatus (o. chrysops as in the proposal),
hybridizing with it, then it follows bairdi
should be considered the second parental species. However, the proposal
includes (from section D and E) that bairdi
is characterized by a stable phenotype throughout central and southwestern
Ecuador, that the southern limit of griseonuchus
is the Ecuador-Peru boundary, and that both forms are reciprocally diagnosable
with no evidence of hybridization between them. A final note to this section is
the most undebatable fact: we need appropriate morphological and molecular
sampling, both in size and geographical coverage, to have a clear answer to the
question about the specific/subspecific status of these forms.
“First,
I think the phenotype of these forms from central Ecuador to northern Peru is
not stable but variable. You can see from a cursory view of different pictures
from their geographical range (Figure 1 below) that there are not clear cut
differences in the extent and coloration of the crown patch and, most
importantly, in the presence or extension of the black nape behind the crown
patch. In fact, Céspedes-Arias et al.
(2021) highlighted the presence of individuals, in their dataset, that could be
considered one form or the other, suggesting that the geographical limits
between both forms are not clear (if in fact there is one limit) and that these
forms, as actually defined, are not clearly diagnosable. Such difficulty to
separate individuals from these taxa were also faced by Chapman (1927) and
Zimmer (1949), but the easier diagnosis as we move toward northern Peru
convinced them to describe and keep this form as a valid subspecies. The
molecular data further complicate things. There are shared haplotypes between
both bairdi and griseonuchus, and
the geographical distribution of those haplotypes is important to consider. The
type locality of bairdi is Cicalpa
Viejo, in Chimborazo, Ecuador, just south of Tungurahua. On the east and
southeast, you can find Sangay National Park and the Cordillera of Cutucú, in
the Morona-Santiago province (localities sampled by Céspedes-Arias et al. 2021). In Tungurahua, the last
locality considered for the hybrid zone, as representative of the parental
phenotype in the south, all haplotypes were either shared with hybrids or
closely related to the ornatus-melanocephalus
haplotype group. From here to the boundary between Ecuador and Peru, bairdi phenotypes south of the type
locality were either genetically closer to ornatus-melanocephalus
or griseonuchus, or hybrid
haplotypes. Similarly, phenotypes in the bairdi
potential range but with some phenotypical similarities to griseonuchus, were closer either to the ornatus-melanocephalus or to the griseonuchus group. This pattern is specifically relevant in the
Morona-Santiago localities where haplotypes shared with griseonuchus were geographically north of those shared with the ornatus-melanocephalus group. Thus,
available data suggest an area of potential genetic exchange/mixture between
the north and the south, resulting in a potential “pure bairdi” geographical range unclear or potentially extremely small.
We could even argue, based on genetic similarities and north-south plumage
coloration pattern, that griseonuchus
might better reflect the parental phenotype from the hybrid zone (in fact, some
haplotypes of ruficoronatus and
hybrids are more similar to the griseonuchus
haplotype cluster than to the ornatus-melanocephalus
group), but it is just an alternative hypothesis.
“In
summary, these taxa are not clearly diagnosable to me and recognize even their
subspecific status might require a thorough morphological study.
“F.
Recognize black crown phenotypes, south of the Marañon River, as melanocephalus This section recognizes
phenotypic differences in melanocephalus
north and south of the Marañon River (chestnut vs. black crown, respectively),
in correlation with the potential geographical/ecological barrier represented
by this river and associated with many bird Andean splits. The issue here are
the haplotypes found in the only malaris
population studied (Amazonas Department, Peru), which are shared both with griseonuchus and melanocephalus/bolivianus.
Molecular divergence from the mtDNA (ND2) is just 0.5%, and a major difference between
malaris and melanocephalus/bolivianus
(malar stripe connecting the lores and auriculars and interrupting the yellow
eyering) is shared between griseonuchus
and malaris (Zimmer 1949). This same
author found two individuals (of malaris)
with traces of brown on the base of crown feathers, suggesting intermediacy
between these forms. Something interesting I noticed, looking at pictures from
the northern hybrid zone, is that some hybrid birds approach the black crown
phenotype of the south, suggesting it might not take much to switch from one
phenotype to the other (Figure 4 below).
“Figure 4. Variation of plumage coloration in Myioborus melanocephalus south of the
Marañon River (Peru and Bolivia). Ventral coloration and face patterns show
variation throughout the distribution of these three taxa. The first picture
represents a hybrid bird from Putumayo, Colombia, in which the crown is almost
black, approaching pattern in this group of melanocephalus.
Top row: from left to right, ornatus x melanocephalus, Putumayo, Colombia, https://ebird.org/checklist/S30754385; malaris: Amazonas,
https://ebird.org/checklist/S97463664; Amazonas, https://ebird.org/checklist/S194175525; San Martín, https://ebird.org/checklist/S63751287; melanocephalus: Huánuco, https://ebird.org/checklist/S63850047;
Middle row: from left to right, melanocephalus,
Pasco, https://ebird.org/checklist/S66189166; Pasco,
https://ebird.org/checklist/S215302743; Junin,
http://ebird.org/checklist/S214612265; bolivianus:
Cuzco, https://ebird.org/checklist/S112334054; Puno, https://ebird.org/checklist/S144178912;
Bottom row: from left to right, Cochabamba, https://ebird.org/checklist/S65135514;
Cochabamba, https://ebird.org/checklist/S155269136; Cochabamba, https://ebird.org/checklist/S160415431; Santa Cruz, https://ebird.org/checklist/S198792492”

“G.
Recognize current geographical variation in melanocephalus,
including malaris, melanocephalus and bolivianus. As indicated in this section, these three taxa run
north to south, showing continuous or discrete variation in the characters
defining them, making it difficult to discern geographical limits among them
(Figure 4). Also important to state is that similar haplotypes at either side
of the Marañon River might indicate potential introgression or recent gene flow
suggesting malaris could be
genealogically closer to northern forms (bairdi
and griseonuchus) than to southern
ones (if so, we will need to ask ourselves what the factors are associated with
differences in plumage coloration in these birds). The haplotype network shows
a melanocephalus/bolivianus more cohesive group in comparison to the relationships
of the malaris haplotypes. It is
important to understand that although the Marañon River is an important barrier
for Andean birds, many lineage breaks do not correspond to this barrier
(phenotypically or genetically), and we can have no breaks or finding them
further south (San Martín, Huánuco, Pasco), as shown in phylogeographical
hypothesis for several Andean taxa across the Marañón River barrier (e.g., Mionectes striaticollis, Tangara vassorii, Pyrrhomyias cinnamomeus; Cuervo 2013).
“In summary, regarding the last two sections,
we need thorough morphological studies of variation in the black crown melanocephalus, as well as a better
molecular evaluation of the region in which the crown changes coloration (the
major difference among these taxa). I do not see further splits within melanocephalus, but the case for malaris
is still contentious, which could impact decisions regarding the recognition of
different species north and south of the Marañón River for melanocephalus.
“How
many species in the albifrons-ornatus-melanocephalus
species complex?
“From the evidence available I think M. albifrons is the clearest candidate
for species status. Data suggest this taxon to be the sister species to the
rest of the complex, it seems to be homogeneous both phenotypically and
genetically, and it appears to be isolated from the rest of the complex by the apparent
stronger geographical barrier in the history of this specific group. Its
phenotypic similarity to rufous-crown melanocephalus
might make one to speculate on the potential ancestral phenotype, which could
be present in the northern Andes at least 2-3 Mya, based on the stem age of
this group.
“The case for the taxa north of the
Marañon River is the most complex to understand with the data on hand. We have
a phenotypic hybrid zone that includes a rufous-crown melanocephalus with o.
chrysops, but the mtDNA data show that genotypically the hybrid zone
includes the northern extremes of both ornatus
phenotypes and go farther south of the bairdi
type locality. In fact, Céspedes-Arias et
al. (2021) found phenotypic hybrids farther south of the hybrid zone (but
not as intermediate as you can find in the middle of it). The reality is that
there are still many questions waiting for answers: is the hybrid zone primary
(selective divergence due to environmental conditions or other selective agent)
or secondary (contact between two previously isolated species)?; is it
asymmetric (one taxa displacing the other)? is it advancing or controlled by
selection? We need genomic data, and we could ask how such data could impact
our decisions here. It could indicate that genomic divergence is large among
the taxa involved in the hybrid zone and correlated with phenotypic
differences; if so, we could suggest secondary contact and potential
introgression or positive selection (e.g., selective sweep) of mtDNA, a pattern
misleading the interpretation of the evolutionary history of the complex. On
the other hand, it could show the same mtDNA pattern, suggesting recent
expansion and diversification (as the high haplotype diversity and low
nucleotide diversity could indicate) and a lack of reproductive isolation
between these taxa. Alternatively, we could find lack of general genome
divergence except for few genes that could be involved in plumage color/pattern
differences, suggesting these traits could be under selection. Understanding
these processes is important because it could suggest the hybrid zone is
advancing and homogenizing all populations, or, for example, there is selection
against hybrids and assortative mating occurs and prevents the hybrid zone for
advancing. For me, knowing these patterns/processes will provide the clues to
suggest there are one or several species involved here.
“The case for the black-crown melanocephalus south of the Marañon
River is perhaps less complicated, but the truth is that we know less here.
Morphological variation is far from clear (similar to the case of the
rufous-crown melanocephalus), and we
need thorough studies of geographical variation, both in the phenotype and
genotype. There has been consensus that the three forms south of the river
should be kept together in one species unless future data suggest otherwise.
What is contentious is the case malaris,
as its haplotypes are shared both south and north of the river. I would like to
include an example from the same genus to think about it. In a previous study, M. castaneocapillus was found to be
paraphyletic in relation to M. cardonai,
two species from the Pantepui Region (Pérez-Emán 2005). One subspecies of castaneocapillus was found sister to cardonai, a surprising result as we
thought both cardonai and albifacies were sister taxa (more
similar phenotypically). One possibility would be this pattern reflects
accurate phylogenetic relationships leading to recognize more species (many of
these taxa are geographically isolated) or to invoke introgression (due to some
phenotypic similarities with one unsampled populations of castaneocapillus). A recent genomic study (Zhao et al. 2025) found cardonai sister to albifacies with high support, contrary to our previous study, which
was also backed up with high support (mtDNA). Why do I mention this? Different
strongly supported data could tell us different stories. Available data from malaris and our logical perception that
different phenotypes (rufous vs. black crowns) should be different, suggest the
potential for mtDNA introgression in the range of malaris or incomplete lineage sorting from a recent divergence
potentially correlated with the geographical barrier represented by the Marañon
River. If we split now, what if genomic data suggest both rufous and black
crowns melanocephalus are closer to
each other and both to ornatus? It
could drastically change our perception of species limits here and will promote
further discussion.
“In
summary, I think each of the potential scenarios to consider from this proposal
regarding the number of species included in this complex, from 2 to 6 species,
has merits and are based on different aspects of the data we have available
right now. However, as Céspedes-Arias et al. (2021) concluded, their study
“..provides a starting point for additional research on the dynamics of this Myioborus hybrid zone”. As such, more
data are required to safely make taxonomic moves that are stable on time and
adjust to the established working criteria (species concepts). Time and new
data will get us closer to understand the evolutionary dynamics of this group
and support (or potentially reject) the claims for each of the potential
splitting scenarios. For me, if we are going to make a change to the current
taxonomy, I will stand for a conservative one: two species, M. albifrons and the rest (ornatus, I believe, for priority), with
the reasons clearly stated before. Any move, even with their merits, includes a
certain amount of uncertainty that invites for more changes in the future. The
good news is that Laura (Céspedes-Arias) is currently working on the genomic
characterization of this complex and will likely place us in a more suitable
position to make data supported taxonomic decisions.
“References:
· Céspedes-Arias, L et
al. 2021. Extensive hybridization between two Andean warbler species with
shallow genetic divergence. Ornithology 138: 1-28.
· Chapman, FM. 1927.
Descriptions of new birds from northwestern Peru and western Colombia. American
Museum Novitates 250: 1-7.
· Cuervo, AM. 2013.
Evolutionary assembly of the Neotropical montane avifauna. LSU Doctoral
Dissertations, 275.
· Cuervo, AM & L
Céspedes-Arias. 2023. The type of Setophaga ruficoronata is a hybrid:
implications for the taxonomy of Myioborus warblers. Zootaxa 5383:
476-490.
· del Hoyo, J & NJ
Collar. 2016. HBW and BirdLife International Illustrated Checklist of the Birds
of the World. Volume 2: Passerines. Lynx Edicions, Barcelona.
· Lovette, IJ et al.
2010. A comprehensive multilocus phylogeny for the wood-warblers and a revised
classification of the Parulidae (Aves). Molecular Phylogenetics and Evolution
57: 753-770.
· Pérez-Emán, JL. 2005.
Molecular phylogenetics and biogeography of the Neotropical redstarts (Myioborus; Aves, Parulinae). Molecular
Phylogenetics and Evolution 37: 511-528.
· Tobias, JA et al. 2010. Quantitative criteria for
species delimitation. Ibis 152: 724-746.
· Zhao, M et al. 2025. A phylogenomic tree of
wood-warblers (Aves: Parulidae): Dealing with good, bad, and ugly samples.
Molecular Phylogenetics and Evolution 202: 108235.
· Zimmer, JT. 1949.
Studies of Peruvian birds. No. 54. The families Catamblyrhynchidae and
Parulidae. American Museum Novitates 1428: 1-59.
Additional comments from Areta: “I agree with Jorge´s
in-depth analysis which has added key data and perspectives in an exemplary
analysis (¡gracias Jorgito!). We seem to be largely
aligned in how we see the case, and Jorge´s deep-dive into the case has shed
new light into this. SACC currently considers three species in the complex: albifrons,
ornatus, and melanocephalus. I have expressed my reservations on
whether the split between ornatus and melanocephalus across the
Marañón will hold once genomic and more morphological data are available, as
current data indicate some type of leakage over the valley (but we do not know
in detail what is going on here). Thus, I think that the conservative stance
here would be to keep recognizing three species (albifrons, ornatus,
and melanocephalus) instead of two (albifrons north of the
Táchira depression and ornatus everything south of the Táchira
depression). I am really eager to read Laura´s next work on the complex."
Comments from Robbins: “Impressive in-depth analysis by
Jorge! I think his conservative,
well-reasoned suggestion of recognizing two species, albifrons and ornatus
(apparently that has priority for this group of taxa), is the best course of
taxonomy at this point. Naturally, we
all look forward to Laura Céspedes-Arias genetic data that may provide further
clarification of this complex.
“Major kudos to Jorge for helping sort through things as they now
stand.”
Comments from Claramunt: “I vote conservatively, as the
new evidence does not challenge the current SACC taxonomy and the proposals for
alternative species limits are not backed by published evidence.
“A. YES. Continue
to recognize M. albifrons as a separate species.
“B. NO to
recognize M. chrysops as a distinct species from M. ornatus. It’s
an interesting possibility but there is no new data to support it.
“C. YES to
suppress the name ruficoronatus; eliminate it from being a valid
subspecies.
“D. YES but
not sure how this is pertinent to SACC.
“E. NO.
There is no new published evidence supporting species status for bairdi.
“F. YES to
continue recognizing the remaining southern members of the complex as a
separate species.
“G. NO. As
shown by Cuervo and Céspedes, subdivisions withing the southern group are not
well characterized. I think it’s better to recognize the groups as a single
taxon.”
Comments from Lane: “Thanks for the trees, although
they are "just" mtDNA, so taken with a pinch of salt. For example, I
am floored that M. torquatus is not sister to the remaining M.
melanocephalus/ornatus complex in the Lovette tree, as it certainly
has the voice and plumage for it! Happily, it seems to be sister in the Pérez-Emán
tree. Anyway, here are my votes:
“A) YES to
continuing to recognize M. albifrons as a species.
“B) NO. At
least in the Lovette tree, it appears as though chrysops and ornatus
are interdigitated (but see my comment about M. torquatus above with
respect to the branches on that tree!). Nevertheless, even if these taxa are
allopatric, they seem pretty close in many respects.
“C) YES to
suppressing ruficoronatus.
“D) YES to
recognizing the name bairdi.
“E) NO to
recognizing griseonuchus as a separate species.
“F) YES to
recognizing the birds south of the Maranon as M. melanocephalus, a
separate species from M. ornatus.
“G) A very
weak YES to continue recognizing the subspecies under melanocephalus.
These will likely prove to be clinal, and thus may have no real meaning, but
until that is shown, I am fine maintaining them as named taxa for now.”
Comments from Remsen: “I can’t add anything that hasn’t
already been said in the many good comments above. Thanks especially to Jorge for the helpful,
extensive comments. We currently do not
provide a classification at the subspecies level, but that will happen
eventually, so those parts of the proposal lay some groundwork for that.
“A. YES. No change to status quo warranted.
“B. NO. More data needed.
“C. YES.
“D. YES – insufficient data to make a change.
“E. NO - insufficient data to make a change.
“F. YES - insufficient data to make a change.
“G. YES. Any change is
premature without a quantitative analysis of geographic variation.”
________________________________________________________________________________________
Proposal (1003) to South American Classification Committee
Species limits the Myioborus melanocephalus
complex: Taxonomic options for resolving the classification of the species
forming an extensive hybrid zone between southern Colombia and northern
Ecuador, and related taxa
Antecedentes: Céspedes-Arias et al. (2021) described
in detail an extensive hybrid zone
between a northern taxon, Myioborus ornatus chrysops, and a southern
taxon (M. melanocephalus ruficoronatus (the northernmost subspecies of M.
melanocephalus). This hybrid zone is exceptionally long, covering ca. 200
km between southwestern Colombia and northwestern Ecuador and is apparently
stable (no “pure” parental phenotypes occurring within the central hybrid
zone). They found that the mitochondrial genetic differences along this zone
were extremely small but the phenotypic differences, particularly in plumages
of the head, were visually striking. Treating
the entire melanocephalus complex as a cline, apparent gene flow
along the occurs with only a distinct a break at the dry Marañon valley of Peru
(a well-known barrier separating highland wet-forest taxa on either side).
The taxonomy
underlying the aforementioned study was effectively relegated to the dustbin by
Cuervo & Céspedes-Arias (2023) when they examined the photographs, original
illustrations and description of the type specimen of ruficoronatus and
found that its plumage was identical to those of hybrid specimens from near the
midpoint of the hybrid zone, with its probable provenance from the area of
Pasto, Nariño, Colombia. Here is he
plate from Cuervo and Céspedes-Arias:

This effectively
excludes ruficoronatus as the name of any valid taxon, and
prompted a more far-reaching examination of other taxa, including their type
descriptions, photographs of same, and distributions, initially to find the
“pure” species-level taxon that would represent the southern parent of the
hybrid zone. For brevity here, I omit details of type descriptions and
photographs, for which see Cuervo & Céspedes-Arias (2023). The appropriate
southern parental taxon found was bairdi once the confusion regarding
its distribution and erroneous synonymization under ruficoronatus had
been cleared up. This taxon is found through central and southern Ecuador. They
then proceeded to examine the distributions, type specimens and distributions
of the other taxa of the clade that includes the hybrid zone, and including M.
albifrons of Venezuela, the sister and near outlier to this clade. North of
the hybrid zone, three taxa occur: M. o. ornatus of the Eastern
Andes and M. o. chrysops of the Central Andes of Colombia, as well as albifrons.
Proceeding south on the eastern slope of the Andes, the taxa include griseonucha
(currently considered a subspecies of bairdi), malaris,
melanocephalus and bolivianus (the last three considered subspecies
of melanocephalus). This chain of taxa is bisected by the dry
Marañón valley of eastern Peru. Of the aforementioned taxa, bairdi and
griseonucha occur to the north and west of this barrier, with the three
taxa of melanocephalus to the south and east.
Here is the
distribution map from Cuervo & Céspedes-Arias (2023):
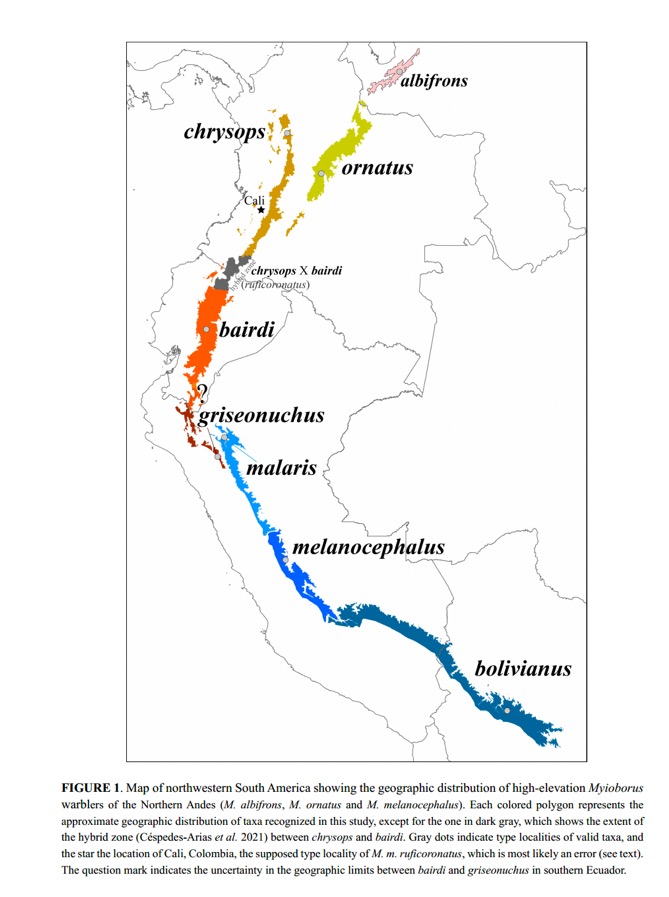
The authors found
that genetic variation over the entire complex permitted recognition of four
genetic clusters, largely correlated with geography and partly with phenotypic
patterns: 1) M. albifrons,
isolated from the remining taxa in the Venezuelan Andes; 2) M. melanocephalus
(including malaris and bolivianus), east and south of the
Marañón; 3) M. bairdi and griseonucha, north and west of
the Marañón, and 4) M. o. ornatus and M. o. chrysops, north of
the hybrid zone. Hybrid genotypes and phenotypes clustered with groups 3 and 4.
They then proposed three taxonomic hypotheses for the species represented among
these taxa:
A. Three species: M. albifrons, M. ornatus
(all of the taxa north of the Marañon and including the hybrid zone) and M.
melanocephalus, south of the Marañón. Thus, ornatus includes a variable
mixture of yellow- and white-faced and partly to entirely black to
rufous-crowned taxa; albifrons has a white eyering and forehead and
black crown, and melanocephalus includes all black-crowned taxa, and is
excluded from participation in the hybrid zone by the intervening,
rufous-crowned bairdi and griseonuchus. The species status of albifrons
is not affected; in fact, its distribution approaches rather closely to
nominate ornatus from the north, but no hybrids between the two are
known.
B. Five species: M. albifrons, ornatus,
chrysops, bairdi and melanocephalus. This option differs in
splitting chrysops and bairdi from ornatus. This is
advantageous in recognizing the hybrid zone as strictly between chrysops and
bairdi. Both melanocephalus s.s. and ornatus s.s. are
isolated from this zone: the former by bairdi (and griseonuchus)
on the opposite side of the Marañón valley, and ornatus from chrysops
by the wide Magdalena River valley. No phenotypic evidence of recent
hybridization between the latter two exists, and their similarity in
mitochondrial genetics possibly best indicates incomplete lineage sorting
during their separation. Moreover, the temperature increase due to ongoing
climate change further reduces probability of future genetic exchange between ornatus
and chrysops, and some checklists have in fact accorded
separate species status to each. Cuervo & Céspedes-Arias (2023)
favored this option.
C. Six species, by the additional separation of griseonuchus
from bairdi reflecting their phenotypic diagnosability and the
apparent absence of hybrid phenotypes between them. The problem here is that
very few specimens are available from the area of possible contact across the
border between Ecuador and Peru; further collecting and field observations are
needed to evaluate this possible split.
For the SACC, I
recommend voting YES for one of the above three options: if any one receives a
majority of votes, it passes; if no option passes, the voting could be repeated
after eliminating the option least voted.
A. Three species
B. Five species
C. Six species
Literature cited:
Céspedes-Arias, L. et
al. 2021. Extensive hybridization between two Andean warbler species with
shallow genetic divergence. Ornithology 138:1-28.
Cuervo, A. M & L.
Céspedes-Arias. 2023. The type of Setophaga ruficoronata is a hybrid:
implications for the taxonomy of Myioborus warblers. Zootaxa 5383(4):
476-490.
Gary Stiles, June 2024
_______________________________________________________________________________________________________________________________________________
Comments from Del-Rio: “YES for B, although I believe whole genome +
habitat studies would benefit the understanding of the strength of potential
barriers to reproduction between chrysops and bairdi.
In the meantime the five taxa should be treated as species.”
Comments from Areta: “YES for A. The species-limits in this
complex are not easy to sort out given the lack of samples from some key areas
and the seeming ability of taxa to interbreed wherever they meet. What a
fascinating system! Genetic divergences are shallow, and support for many of
the mtDNA relationships is poor, although they make a lot of sense in
geographic and plumage terms. Pérez-Emán (2005) uncovered a close relationship
between "ruficoronatus",
chrysops and ornatus. Vocalizations are
structurally very similar across the geographic range of the ornatus-melanocephalus complex, and this applies to albifrons too: all taxa give a rapid series of tonal notes that
includes some shorter and some longer whistles ascending or descending in
frequency that rise and fall in pitch and speed up and increase in amplitude as
the song progresses. All can also duet.
Given that the quite
different-looking bairdi and chrysops hybridize freely and massively
across 200 km, it seems that there are no barriers to interbreeding and that
these two can be considered as part of the same species. The white-faced ornatus is very similar to chrysops, and if one can use the bairdi-chrysops case as a predictor of what would happen if chrysops meets ornatus, the answer would seem to be massive interbreeding. Do the
eastern population of chrysops meet ornatus somewhere? It seems, based on
the examination of a few photographs, that ornatus
at Sumapaz at the southern end of the range have considerably less white on the
face-sides and on the upper-throat (below the bill) [e.g.,
https://macaulaylibrary.org/asset/151865601;
https://macaulaylibrary.org/asset/192970091] than birds in the northern end
[e.g., https://macaulaylibrary.org/asset/205397261]. This suggests that the
unsampled area between ornatus and
eastern chrysops may well show a
broad area of phenotypic transition.
“The situation
between bairdi and griseonuchus seems relatively
straightforward: the plumage variation reported by Cuervo & Céspedes-Arias
(2023), and the genetic data are consistent with considering them as part of
the same species. Therefore, in my view, also conspecific with chrysops and ornatus.
“The most complicated
part to me is the trans-Marañón divergence between griseonuchus and malaris.
Céspedes-Arias et al. (2021: 12) put it this way: "Genetic structure was
shallow, and only clearly associated with 1 topographic discontinuity of the
cloud forest belt. We found evidence of mtDNA differentiation across the
Marañón River Valley, a dry area that dissects the cloud forest distribution of
M. melanocephalus and coincides with the transition between
rufous-crowned (M. m. griseonuchus) and black-crowned (M. m. malaris)
forms (Zimmer 1949, Curson et al. 1994). This arid valley is important for
differentiation in many other cloud forest birds (Bates and Zink 1994, Chaves
et al. 2011, Gutiérrez- Pinto et al. 2012, Winger and Bates 2015), and likely
acts as a strong barrier for Myioborus taxa restricted to humid
high-elevation forests and scrub (Curson et al. 1994). The exception are 3
specimens from south of the Marañón (Amazonas, Peru), corresponding to the
black-crowned subspecies M. m. malaris, which clustered with individuals
of M. m. griseonuchus and M. m. ruficoronatus occurring north of
the Marañón Valley. This pattern might reflect trans-Marañón introgression
(Winger 2017) or incomplete lineage sorting (Maddison and Knowles 2006)."
Thus, 3 out of 8 samples of the black-crowned malaris from immediately south of the Marañón valley clustered with
the rufous-crowned griseonuchus,
while 5 out of 8 samples of malaris
from the same area clustered with the black-crowned melanocephalus-bolivianus
(see Figures 1A and 2A in Céspedes-Arias et al. 2021). This is only mtDNA data,
but the fact that there is trans-Marañón mtDNA sharing does not instill
confidence on the degree of isolation between the similar (yet differently
crowned) griseonuchus and malaris: if there is leakage across this
barrier, these two sets of taxa would very likely interbreed freely if given a
chance to do so. Here is when having genomic data would provide us with much
needed insights to inform our taxonomic decisions.
“I think that the
most conservative course of action in the light of the great works by C-A et
al. 2021 and C & C-A 2023 is to vote for their "three-species alternative" (Option A). Maybe a genomic
dataset can make my understanding change and result in more splits, but at
present, I think that even the three-species treatment might be on the generous
side of splitting.”
Comments from Robbins: “Of the options given, I vote for A. Given the data at hand, I believe Nacho’s
interpretation of how best to treat these taxa is the best option. Another
option that hasn’t been given is a four species treatment, i.e., albifrons,
ornatus, chrysops, and melanocephalus. By doing the latter,
one does not have to speculate on what might happen if ornatus and
eastern chrysops should be found
in contact. Nonetheless, I find
Nacho’s observation that there may be a cline in white in ornatus or may
even indicate hybridization with eastern chrysops to be plausible. So,
if four species treatment isn’t an option, I vote for option A. Clearly, more
study is needed on what is going on in the Marañón
region with regard to griseonuchus and malaris.”
Comments from
Jaramillo: “YES on B – I can
see both the 3 and 5 species solutions as valid given what we know now. When
the genomic analysis happens, if ever, we will know more. But to me the 5
species solution seems cleaner to me actually. Although super on the fence
here.
Comments from Stiles: “B – 5 species. Here, I comment on Nacho’s conclusion that M.
ornatus s.s. and M. chrysops would show massive interbreeding should
they come into contact, because this has occurred between chrysops and bairdi,
given the shallow genetic divergence in both cases. However, the use of the
latter as a yardstick for predicting the former ignores the complex Andean
topography. The long hybrid zone between chrysops and bairdi occurs
along a continuous range of upper temperate-zone forest habitat, north of which
chrysops occurs alone along the Central Andes at similar elevations for
several hundred km more. The distribution of ornatus s.s. lies between
the northern end of the Cordillera Oriental south to Sumapaz massif. South of
this massif, this cordillera narrows abruptly to a long ridge separated from
the Cordillera Central by the upper Magdalena Valley with tropical elevations
of ca. 350-400m as far south as the latitude of San Agustín. Southward, this
cordillera connects with the Cordillera Central via an eastward extension of
temperate-zone habitat in southeastern Cauca – western Putumayo. The important
point here is that the long ridge south of the Sumapaz massif lowers to
subtropical elevations over much of its length, where the only Myioborus present
is miniatus. At the temperate-zone connection to the south, only chrysops
is found along the highest ridge of this zone – which adjoins the hybrid
zone with bairdi. To the north, the two main cordilleras are separated
by the lower and middle Magdalena Valley which here supports a zone of hot, dry
to moist tropical forest at elevations of 100-200m, over a 50-100+ km wide
zone. Hence, there is NO point of direct contact between chrysops and ornatus
s.s. anywhere in their respective distributions – they are isolated at
least since the latest glacial maximum and given the warming trend of
climatic change, this degree of isolation can
only increase in the coming years.”
Comments from
Bonaccorso: “YES for A. Based
on the available evidence, three species (Myioborus albifrons, M. ornatus, and M. melanocephalus) make sense
(five also, but I think the move would be too bold). Céspedes et al. are
working on a genomic treatment of the whole group, and knowing Laura
(Céspedes), it will be superb. So, we will have more than enough genetic
evidence to consider further splits.”
Comments from Lane: “I am not entirely clear of the accepted
phylogeny within this clade, but the evidence provided makes me think that
there are a couple of other taxonomic options that are not offered here
(namely, 2 species [M. albifrons and M. melanocephalus, including
all remaining taxa of the complex] and 4 species. The one I would favor, based
on what I am seeing here is: 4 species.
1. M. albifrons (monotypic)
2. M. ornatus (monotypic)
3. M. chrysops (assuming that it is
decided to be the name with priority over M. bairdi, and also including M.
griseonuchus. The broad hybrid swarm zone seems to make this a necessity)
4. M. melanocephalus (including malaris
and bolivianus).
“Did I miss some
reasoning why this taxonomy was not considered an option? So, NO for any of the
three options offered in the proposal.”
Additional comments
from Areta: “Thanks Gary for
clarifying that the gap between ornatus and chrysops is real. In no way have I ignored the
complex Andean topography in my reasoning. However, the meager plumage, vocal,
and genetic differences that we know at present, leave ornata as a very weak species in my perspective,
and I still stand by the yardstick approach that it is better considered a
subspecies of chrysops. It is
great to hear that Laura is working on a genomic perspective on these Myioborus. I am eager to see what
she finds, and to revise my vote accordingly.”
Comments from
Claramunt: “YES to A. Three
species. The new information clearly shows gene flow and introgression between chrysops and bairdi, demonstrating that
they are not separate species. Not much we can say about the rest of the
complex, but I agree in maintaining albifrons and melanocephalus as separate species for now.”