Proposal (1009) to South
American Classification Committee
Treat Amazona
autumnalis (Red-lored Parrot) as consisting of three species
Note:
This is a high-priority issue for WGAC.
Background: Our current Note reads as follows:
34c. The Ecuadorian subspecies lilacina
and the Brazilian subspecies diadema were formerly (e.g., Cory 1918, Pinto 1937) considered separate species
from Amazona autumnalis, but Peters (1937) treated them as
conspecific. Ridgely & Greenfield (2001) treated diadema as a
separate species but did not provide justification. Del Hoyo & Collar (2014) treated both lilacina (“Lilacine
Amazon”) and diadema (“Diademed Amazon”) as separate species based on
differences in plumage and bare parts coloration, and this was further
supported by qualitative inspection of specimens by Donegan et al. (2016). Smith et al. (2023) found a deep genetic
divergence between autumnalis s.s. and diadema. SACC proposal needed.
The
status quo, maintained in most classifications (e.g. Dickinson & Remsen
2013), is that this species consists of 4 subspecies:
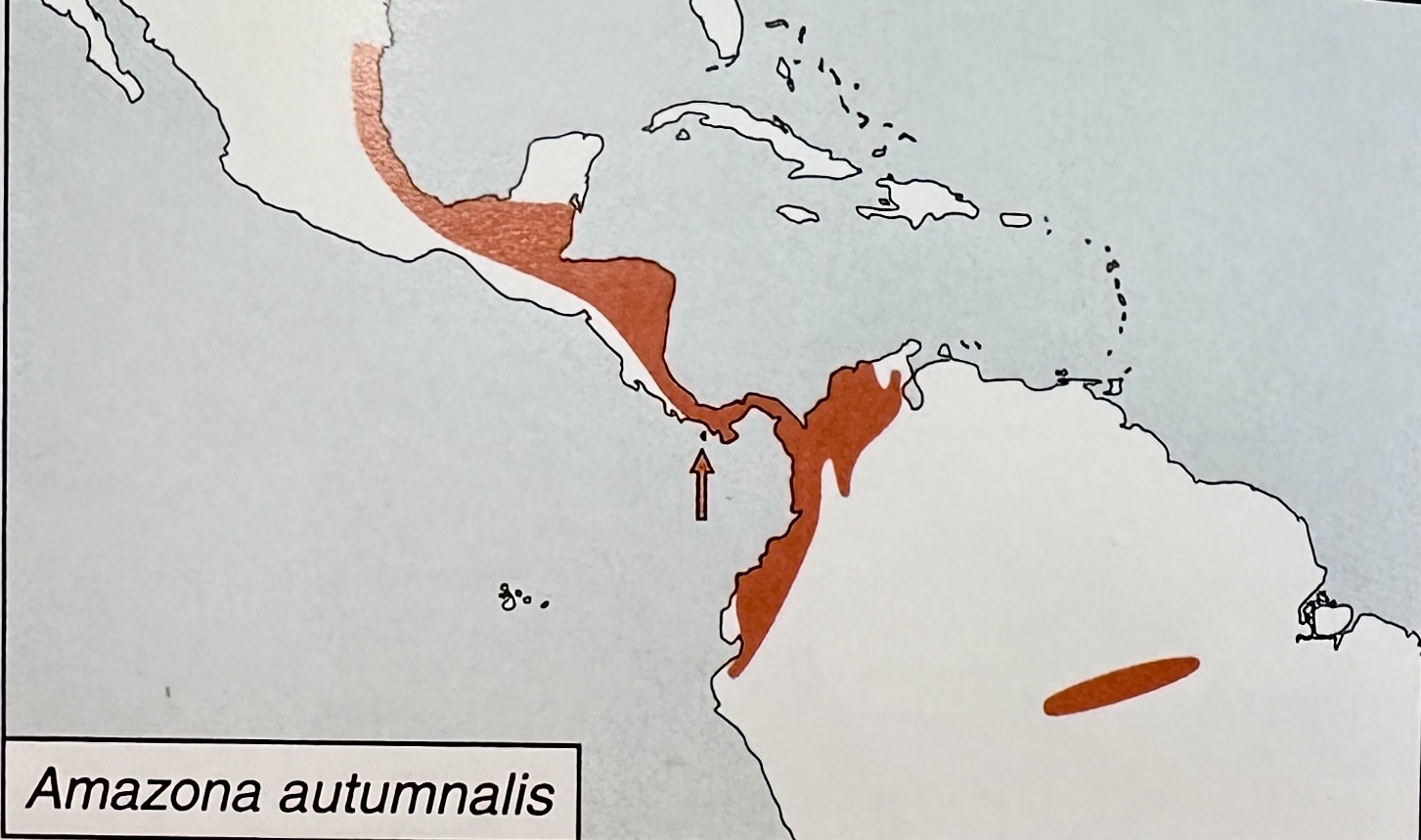
1. A. a. autumnalis from Mexico to N.
Nicaragua
2. A. a. salvini from Nicaragua to the
Chocó of sw. Colombia and also to n. Venezuela.
3. A. l. lilacina: endemic to the Chocó
of w. Ecuador
4. A. a. diadema: disjunctly in central Amazonian
Brazil along the Amazon and lower Rio Negro.
Here
is the composite distribution map from Forshaw (2006:”Parrots of the World”):
Here
is the plate by Frank Night from Forshaw (2006:”Parrots of the World”):
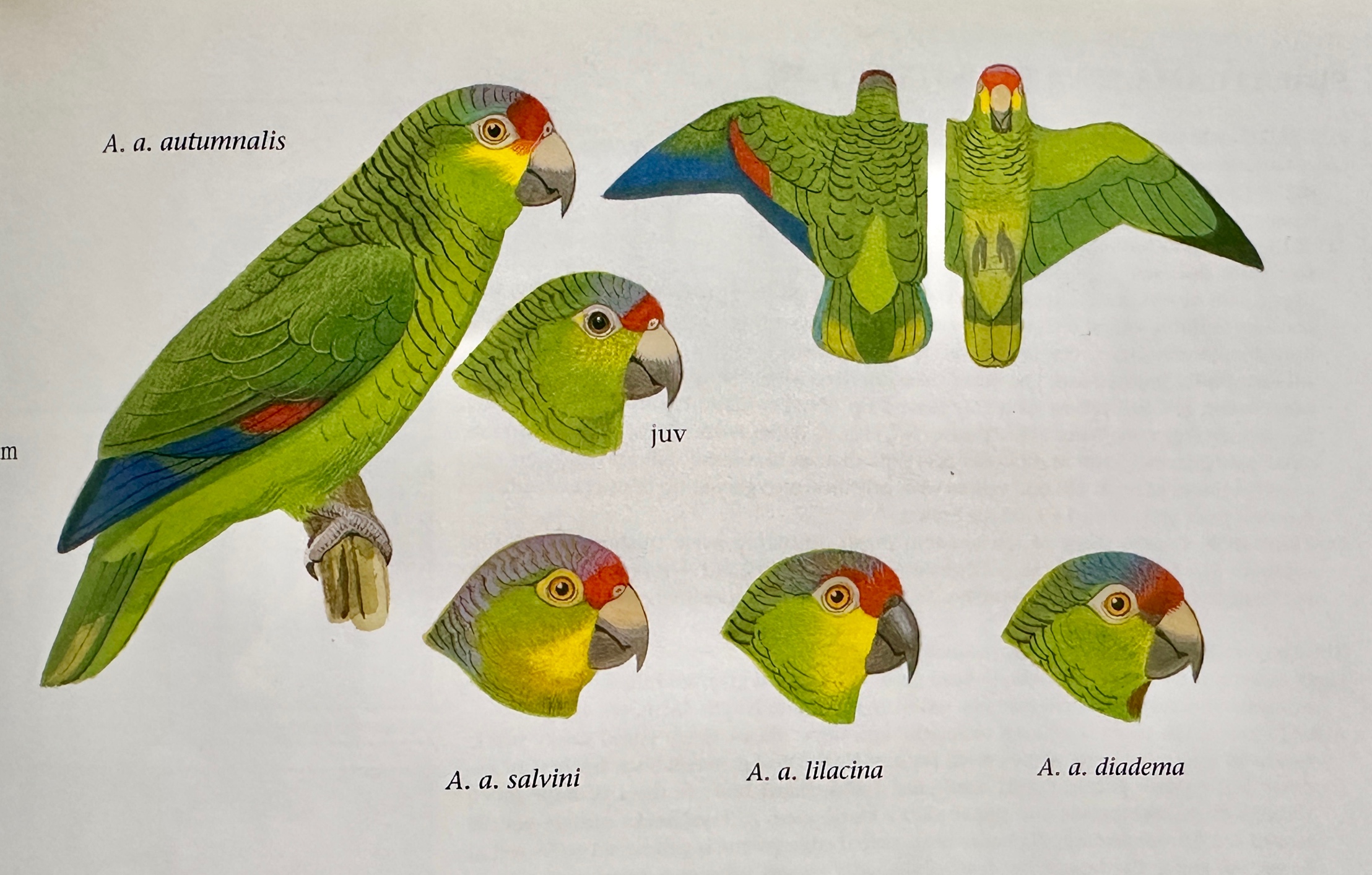
A.
l. salvini
apparently has a zone of intergradation with A. a. autumnalis.
Here’s
the plate in del Hoyo & Collar (2014) by Ångels Julglar:

Here
are three photos from Macaulay, L to R, nominate autumnalis pair (by
John Doty), lilacina (by Edison Buenaño), diadema
(by Héctor Bottai):

New
information:
Del Hoyo & Collar (2014) treated lilacina
and diadema as species separate from autumnalis. Here is the HBW rationale (provided by Pam
Rasmussen):
For
diadema:
“Usually
considered conspecific with A. autumnalis and A. lilacina, but
differs from lilacina in certain characters outlined under that species, and
from both in its nares being covered in red feathers, with rest of red patch on
face sharply delineated to form a distinct rectangle, quite different from
shape of red patches on other taxa in complex, and allowing blue on crown to
extend forward to border the frons (3); powdery dorsum, resembling more that of
A. farinosa (1); bill black but with pale patch below nares (2); head
much flatter and longer in lateral profile (mensural score 1). Monotypic.”
For
lilacina:
“Usually
considered conspecific with A. autumnalis and A. diadema, but
differs in its all-black upper mandible (2); red of forecrown continuing over
eye (2); lilac of crown not extending onto nape (2); paler, clearer green cheek
(ns[1]); possibly no red on chin (ns[1]); narrow, sharp yellow edges of
wing-coverts and flight-feathers (ns[1]); retiring, non-aggressive behaviour (quite different at least from autumnalis (1) )
(ns[1]); smaller bill (tiny museum sample size supplemented by photographs; 1).
Specific status considered appropriate also in unpublished recent taxonomic
study (2). Monotypic.”
Donegan
et al. (2016) evaluated the del Hoyo & Collar taxonomy with respect to how
this affected the Colombian bird list.
They also provided photos of 5 specimens
from AMNH. Here is what they wrote:
“We concur that A. lilacina is
a quite different bird from both diadema and autumnalis (Fig.
15). It has virtually no blue on the crown but extensive yellowish-green
plumage on the face and throat, carpal edgings and vent and a dark bill.
“The split of diadema from autumnalis
is less clear-cut. The more extensive blue scaling on the mantle of diadema
is a noteworthy feature (Fig. 15), but the head patterning and overall
coloration is more similar to autumnalis. The difference in nare feathering can be seen in online photographs of live
birds but was not obvious in specimens we inspected. Differences in the shape
of the red patch on the lores are less impressive when one considers variation
in this feature both within autumnalis and between the nominate
subspecies and salvini. It is not necessary for us to express any view
on the rank of diadema, as it is extralimital as regards Colombia.”
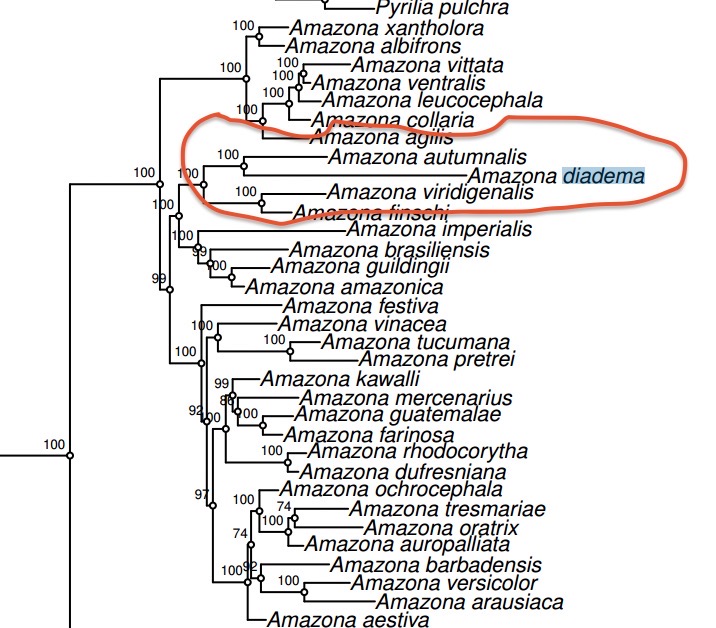
Smith
et al. (2023) did not sample lilacina, but they had diadema in
their tree. It comes out as sister to autumnalis
in their analyses, but the divergence is deeper than that among many taxa
ranked as species in Amazona. In
S27, my eyeball extrapolation down to the X-axis suggests the node that joins autumnalis
and diadema is ca. 3 MYA.



Voting: Let’s do it this way:
YES = Treat lilacina and diadema as
separate species from A. autumnalis.
NO = maintain status quo OR treat either lilacina
or diadema as a separate species but not both.
Discussion
and recommendation:
I’m not going to vote until I see what others say, but I’m leaning towards a
YES for the following reasons. First,
that deep genetic split is impressive. I
don’t support using genetic distance of neutral loci as a metric for assigning
species rank, but this looks like an extreme case, with diadema more
divergent from autumnalis than are many sister taxa treated as species
in Amazona. As for lilacina,
available evidence suggest an abrupt change from A. a. salvini in
Colombia to lilacina in Ecuador, or perhaps this should be phrased to
indicate that there is currently no evidence for gene flow in these parapatric
taxa. If that is really the case, then
species rank is required. Finally,
Peters provided no rationale for the lump, nor has anyone subsequently.
However, here’s what makes me
wonder. The large distance between diadema
and the closest autumnalis populations suggests a long period of
isolation, unlike many sister parrot species, so perhaps that genetic distance
is just the long accumulation of “irrelevant” neutral mutations. The absence of genetic samples of lilacina
is unfortunate – that’s a case in which genetic data really would illuminate
species limits because of the apparent parapatry: other than some governmental
border posts, there is no substantial barrier to gene flow between the
two. As noted by Donegan et al. (2016)
an argument can be made that diadema looks more like autumnalis
than lilacina does. Also, what is
really known about the potential contact zone between autumnalis and lilacina? There do not seem to be many specimens of lilacina
in collections, but it looks to me as if careful photographic sampling of
individuals might hold promise for at least a preliminary assessment of the
contact zone using bill color and head pattern.
As for plumage …. I’m still trying to
reconcile why the specimens shown in Donegan et al. show what I would consider
“important” differences, as reflected in del Hoyo & Collar’s description,
in contrast to my impressions from the photos of living birds, the signal from
which I perceive is dominated by similarities, as in the two plates. Also note the differences between the two
individual autumnalis in the photos – this is almost certainly a mated
pair. How much individual variation is
there? So, I don’t know what to make of
the differences in head plumage. All
this makes me want to demand an actual formal analysis of plumage characters,
with specimen N and locality data reported, before taking the plumage into account. Evaluating N=1 samples of specimen photos,
live bird photos, and artists plates is not the way science should be
conducted. Finally, as for Peters, those
of us who use the Cory-Hellmayr series know that Cory’s early, single-authored
volumes (1918, 1919) made few changes in taxon rank
from the original ODs unless Ridgway or someone else had already done it; not
until Hellmayr joined the project was more critical thinking applied to lumps
and splits.
And then there is the absence of vocal
data. This is tough to analyze in
parrots for obvious reasons, but at the least flight calls seem to be fairly
stereotyped and a good marker for species differences in sympatric congeners.
English
names: If the proposal passes, then I see no need
for a separate proposal on English names given that Lilacine
and Diademed have been used extensively (Diademed even goes back to Cory
1918). If anyone objects, please speak
out.
References:
DONEGAN, T., J. C. VERHELST, T. ELLERY, O. CORTES-HERRERA, AND P.
SALAMAN. 2016. Revision of the status of bird species
occurring or reported in Colombia 2016 and assessment of BirdLife
International's new parrot taxonomy. Conservación Colombiana 24: 12–36.
SMITH, B. T., J. MERWIN, K. L. PROVOST, G. THOM, R.
T. BRUMFIELD, M. FERREIRA, W. M. MAUCK III, R. G. MOYLE, T. F. WRIGHT, AND L.
JOSEPH. 2023. Phylogenomic analysis of the parrots of the
world distinguishes artifactual from biological sources of gene tree
discordance. Systematic Biology 72: 228-241.
Van Remsen, June 2024
Comments
from Robbins:
“NO. Because of the lack of genetic data for lilacina coupled with what
Van points out as potential variation in head pattern (plumage, soft part
coloration), for now I vote NO until some of these issues are addressed.”
Comments
from Jaramillo:
“YES – I tis a patchwork of information here, that I am using. First large
divergence time with diadema based on molecular data. Then lilacina
is quite different in appearance and appears to be essentially parapatric with autumnalis.
That is interesting. I tis piecemeal, and patchwork but I think the three species
theory will be what will prevail over time, as more data is gathered.”
Comments from Areta: “I vote NO to the split
of lilacina. I am unimpressed by the
differences between the different populations in terms of genetic and plumage,
which seem on par with those between several populations of other Amazona taxa.
For example, I myself have studied and published on the sierran populations of Amazona aestiva, which lack any red or
yellow in the "shoulder", no yellow on the face, and a different
green colour despite the close proximity to typical xanthopteryx populations (Areta 2007).
“Terry
Chesser suggested that the deep branch between lilacina and autumnalis
in Smith et al. might be due to missing data (as these taxa are in the upper
10% of missing data taxa).
“In
a PhD thesis (Pilgrim 2010, courtesy Paul F. Donald) the genetic data (883bp of
ND2) tells a different story:
"Table 4.2. The average p-distances between the subspecies
|
|
|
1 |
2 |
3 |
4 |
5 |
6 |
|
1 |
A. a. lilacina.
|
|
|
|
|
|
|
|
2 |
A. a. salvini |
0.012 |
|
|
|
|
|
|
3 |
A.a.autumnalis |
0.023 |
0.026 |
|
|
|
|
|
4 |
A.a. diadema |
0.011 |
0.000 |
0.025 |
|
|
|
|
5 |
A. viridigenalis |
0.053 |
0.026 |
0.057 |
0.057 |
|
|
|
6 |
A.leucocephala |
0.177 |
0.171 |
0.152 |
0.169 |
0.197 |
|
|
7 |
Pionus
maximiliani |
0.369 |
0.381 |
0.341 |
0.376 |
0.336 |
0.353 |
|
Table 4.2. Estimates of Average Evolutionary
Divergence over Sequence Pairs between Groups. |
“The divergence of A. a. lilacina from A. a.
salvini is 1.2% and divergence of A.
a. lilacina from A. a. autumnalis
is 2.3%.
A. a. diadema shows no divergence from A. a. salvini. The A. a.
autumnalis group shows a divergence from A. viridigenalis of between 2.6 to 5.7% and a divergence from A. leucocephala from Cuba of between
15.2 to 19.7%."
“And using the same molecular ND2 dataset,
Pilgrim obtained the following MP consensus tree:

“Pilgrim also reports on the smaller bill of lilacina, and mentions lack of sexual dimorphism in lilacina (as a difference between the
size-dimorphism in other autumnalis
taxa, and Amazona in general).
“A paragraph on geographic
variation in plumage is worth quoting in full:
"The A. autumnalis specimens from the Natural
History Museum at Tring are arranged taxonomically and by country of origin.
While examining these to examine putative differences in area of plumage color
between the subspecies, it was noted that the amount of yellow on the cheeks of
A. a. autumnalis, the feature which gives this parrot its common name of
Yellow–cheeked Amazon, gradually decreases in size from the northern Mexican
specimens with a very large bright yellow cheek patch, to specimens which have
a very small patch of yellow feathers on the cheeks in the south of its range. A.
a. salvini has no yellow in the cheeks. It was not possible to assign some
specimens that had merely a single fleck of yellow on the cheeks to either A.
a. salvini or A. a. autumnalis. These intermediate specimens
originate from Honduras and Nicaragua, an area between the ranges of A. a.
autumnalis and A. a. salvini (Fig. 1.00), and may be subspecific
hybrids as reported by Howell
(1957). No such
intermediate forms between A. a. lilacina and A. a. salvini have
been observed either in the museum specimens or amongst the dozens of wild
captured living specimens observed during my role as European studbook keeper
for A. a. lilacina. This suggests an absence of a hybrid zone between A. a. lilacina and A. a. salvini. However, a survey of the area where the ranges of
these subspecies meet on the border of Ecuador and Colombia would be required
to ascertain this with any certainty. As mentioned in the introduction Olivares (1957) suggests the possibility
of hybridisation but presents no evidence for it. There are no other or more
recent reports of a hybrid zone between A.
a. lilacina and A. a. salvini in
the literature. A. a. diadema has an
allopatric range."
“Areta, J.I. 2007. A
green-shouldered variant of the Turquoise-fronted Amazon Amazona aestiva from the Sierra de Santa Bárbara, north-west
Argentina. Cotinga 27: 71-73
Pilgrim, M.A., 2010, An
investigation into the taxonomic status of Amazona
autumnalis lilacina using a multidisciplinary approach. PhD thesis,
Liverpool John Moores University, Liverpool, UK.”
Comments
from Stiles:
“NO. The data are just too patchy and inconsistent, and it would seem that more
intrapopulation variation in plumage might exist as well; clearly more
thorough, quantitative data on plumage and genetics are needed.”
Comments
from Lane:
“NO. This is a project or two that might benefit greatly from directed
investigation into the relationships of these three groups of taxa. I suspect
both lilacina and diadema will prove to be species-level with
respect to the autumnalis group, but for now we have missing
information.”
Comments
from Claramunt:
“YES. Going on a limb in this case and I agree that
it would be better to have more detailed analyses but at least lilacina looks like a
well-delineated taxon. I don’t see any signs of intergradation in photos from W
Ecuador. The case of diadema is harder because the bird is remotely allopatric and the
diagnostic traits may be just too subtle, but I think it is correct that the
exact shape of the loral/frontal patch is diagnostic, ending in an abrupt
transition to a pure green supercilium in diadema rather than a
diffusing over the eye like in autumnalis. That, plus
the considerable genetic divergence convinced me that we need to revert the
unexplained lumping made by Peters et al.”
Comments
from Zimmer:
“YES. I will admit up front to not
having any experience with lilacina, so the lack of any genetic data for
the latter taxon is vexing, because it doesn’t leave me a lot to go on. I will say, as others have suggested, that
the variation in the amount of yellow in the face doesn’t impress me either –
other Amazona species (amazonica and, especially, aestiva come
quickly to mind) exhibit tremendous individual variation in the amount and
distribution of yellow on the head and face, even within the same population,
and I suspect at least some of this variation is attributable to age. However, I am struck by the fact that, as Van
states, lilacina and salvini have seemingly parapatric
distributions with respect to one another, with no obvious geographic barrier
separating them, and yet, there’s no obvious intergradation, and the respective
phenotypes seem to be maintained. That,
combined with the fact that this is yet another case of a pair of taxa that
were described as distinct species, and then lumped without rationale provided
by Peters, are enough to tilt me in favor of treating them as distinct
species. As for diadema, I feel
on firmer ground in voting for a split on this one. We do have indication of a deep genetic split
on this one, and even though Van’s caveat regarding the possibility of long
isolation just reflecting the accumulation of irrelevant neutral mutations is
well taken, I’ve always felt that diadema was a different beast. I had a lot of experience with autumnalis
in Mexico, Panama, and Costa Rica before I first encountered diadema on
my first visit to Manaus, and I remember being struck by how different it
looked (particularly the farinosa-like abundant powder-down on the
dorsum and different distribution of red and blue on the head, not to mention
the absence of yellow in the face) and sounded.
To me, the flight calls of diadema and autumnalis are
pretty different. I haven’t heard the
songs of either taxon much in recent years, so I’m a little rusty on those, but
the flight calls are what you hear mostly, and my impression of those has
always been that they are distinct. So
to me, if you’ve got two taxa that differ in some aspects of plumage, differ in
at least some vocal characters, and have a deep genetic split relative to other
recognized species-pairs in the same genus, and are clearly on independent
evolutionary trajectories given their extremely allopatric distributions, it
doesn’t really matter whether the depth of the genetic split is largely due to
accumulation of irrelevant neutral mutations or not – they already look and
sound consistently different, and they aren’t going to be sharing genetic
material anytime in the foreseeable future (= ever), so why not revert to the
pre-Peters treatment as distinct species?”
Comments
from Del-Rio:
“NO: Given the limited genetic data available on lilacina, I believe
that future research could potentially reveal that these are all distinct taxa.
However, for the time being, I will adopt a conservative approach. Again, an
interesting system to track different patterns of accumulation of
psittacofulvin in Psittacidae populations living in different environments.”
Additional
comments from Stiles:
“Given Kevin´s comments and reconsidering the information in the BMNH paragraph
as well as other data mentioned by Pilgrim in particular, I am willing to
change my vote to a YES for recognizing lilacina and diadema as
separate species from autumnalis. The case for diadema seems
especially clear for species status given the deep genetic difference, the
morphological and apparent vocal differences and its widely divergent
distribution. The case for species status of lilacina is somewhat less
clear, but given the differences in plumage and bill colors. overall size and
lack of sexual size dimorphism and the lack of any intermediate specimens (of
many examined) I think that this tips the balance toward at least tentatively
considering species status (especially because Peters did not give any real
evidence for lumping them, The lack of specimens (or observations) in the
potential contact zone could be interpreted simply as missing evidence, but on
the other hand, it could indicate that there IS no contact, with lilacina
thus considered as a peripheral isolate from autumnalis which as such evolved a suite of phenotypic differences
(often the case in such situations).”
Comments
from Remsen:
“YES. After letting this incubate for
months. First, Kevin’s comments are
persuasive, even though the data on flight calls are not published. As for the genetic data, I don’t think we can
rely on the unpublished Pilgrim data, which claim that autumnalis and diadema
are identical at those loci. Given the
vast geographic distance between the two and the plumage differences, I think
something must be amiss with those data, or at least they should be repeated. The data of Smith et al. ARE published, and although the branch length might be
exaggerated by missing data, they are not identical. As for lilacina, yes the contact zone
if there is one is not sampled but I think the absence of any signal of gene
flow with the data at hand puts burden-of-proof on treating them as
conspecific. This is a tough call and a
data-deficient case, for sure, but with considerable trepidation, I think what
we have available suggests separate species.”
Comments
from Mario Cohn-Haft (voting for Bonaccorso): “YES. Full disclosure: I know only diadema
in the field. that makes my personal experience with the complex pretty well
useless. However, when has that kept me from offering an opinion? All my
instincts say that diadema must be a distinct species from the rest:
it's very distantly isolated geographically in the central Amazon from all the
others, which are trans-Andean and more or less continuous in distribution; it
has no yellow on the face but often does have yellow on the crown, both of
which seem intuitively like important differences; also has what strikes me as
a wider, more conspicuous bare eyering than the others, more and different blue
on the crown; and finally it has an unusual and mysterious life-history,
apparently involving an intra-Amazonian migration, such that I'm not sure we
even know where it breeds! See below for
a few pictures of diadema from Manaus. So, if there were no genetic data, I'd vote
for recognizing diadema as a full species and would more or less
instinctively visualize the other 3 as geographic variants of a single species,
considering their seemingly continuous distribution. However, the fact that experienced folks (and
the pictures) suggest that lilacina too is different might convince me
to vote "yes" overall.
“However,
there ARE genetic data. The study showing diadema to be strongly
different from autumnalis jibes with the plumage and biogeographic
arguments laid out above and is intuitively satisfying. The Pilgrim study that
includes lilacina and finds all 4 taxa to be between 1-2.5% different
from one another, except diadema which is genetically identical to salvini,
makes no sense at all, and I'm inclined to follow Van's lead and just assume
it's wrong. Not a very satisfying conclusion, i recognize, but seemingly
advisable given the rest of the evidence.
“I
was about to wimp out and vote "no" because of the incomplete and
conflicting genetic data and in recognition that i don't feel like i can say
anything first-hand about lilacina. But then i reread the intro to
the proposal and remembered that the only reason we're even talking about this
is because of Peters' lumping without explanation (as usual). So, really,
the whole thing is quite simple: ignore Peters and vote YES in
recognition of what was right from the start and never should have been
changed.”