Proposal (1010) to South
American Classification Committee
Treat Cranioleuca
marcapatae weskei as a species
Note:
This is a high-priority issue for WGAC.
Background: I (Remsen 1984) described weskei
as an isolated subspecies of nearby ecological equivalent Cranioleuca marcapatae,
based on their conspicuous differences in crown color. The rationale for subspecies rank was based
in part by extrapolation to the situation in Cranioleuca albiceps, in
which (1) the nominate subspecies shows individual variation in crown color
from white to ochraceous, with white-crowned individuals looking superficially
like weskei (to the point that Vaurie [1980] considered them to
represent an isolated population of C. albiceps, although I regard that
as a lapsus), and (2) the Cochabamba subspecies discolor has no white in
the crown. The basic logic was that if
crown color can vary dramatically within individuals of a single taxon as well
as between taxa currently treated as subspecies, then weskei should also
be treated at the subspecies rank.
Here
is the plate from the OD (by Larry McQueen):
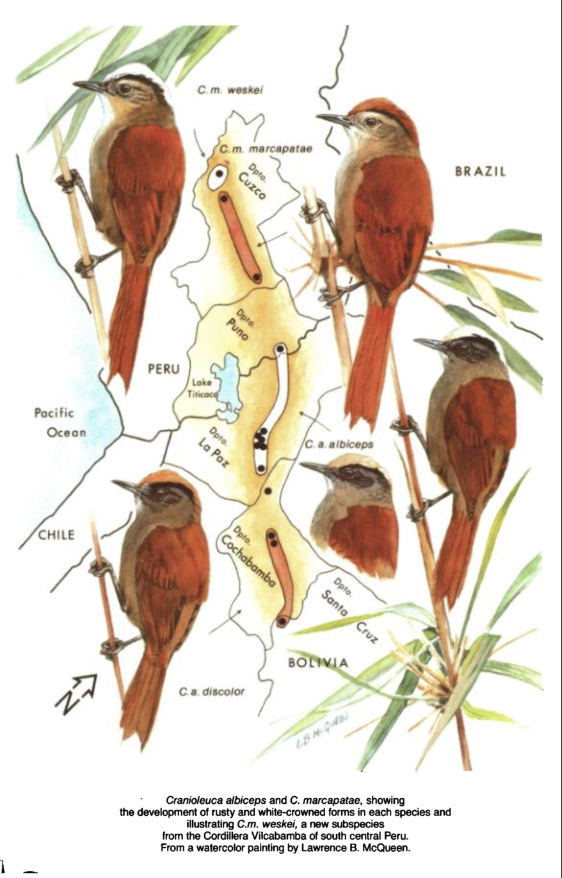
Here
are some specimen from LSUMZ. I don’t
think any of the specimens from dpto. La Paz are as ochraceous as those of C.
a. discolor. Note the crown color of
marcapatae differs from any of the others and matches the back color, unlike
any of the others, including weskei. We don’t know (yet) whether the
tawny crowns in nominate albiceps are a consequence of gene flow:

We
do not have a SACC Note on weskei.
What we do have is the following (which needs to be updated with respect
to this proposal; although I have completed adding SACC notes on del Hoyo &
Collar [2014; nonpasserines], I stalled out somewhere in the suboscines on the
passerine volume [2016]:
49. The superspecies relationship proposed for Cranioleuca marcapatae
and C. albiceps (e.g., Remsen 1984) is corroborated by genetic
data (García-Moreno et al. 1999b, Derryberry et al. 2011); Fjeldså & Krabbe
(1990) proposed that they might be best treated as conspecific.
Here’s
how I set up the taxonomic problems posed by this complex:
“Should
each of the four spinetail taxa in question be considered allospecies of a
superspecies complex, subspecies of a single biological species, or some
intermediate combination? In the absence of comparative data on vocalizations
and other potential reproductive isolating mechanisms or on degree of genetic
differentiation, any taxonomic decisions at this point for these primarily
allopatric populations are tentative at best.”
Concerning
the crown color differences, here is what I wrote:
“Current
information indicates that crown color differences do not necessarily result in
reproductive isolation. Specimens from an area (Choro, Prov. Ayopaya, Dpto.
Cochabamba) geographically intermediate between populations of C. a.
albiceps and C. a. discolor are intermediate in crown color between
the white-crowned nominate subspecies and tawny-crowned discolor.
Furthermore, the high frequency of buff-crowned individuals in the Dpto. La Paz
population (see below) may indicate gene flow from the southern populations.
Thus it seems best to disregard crown color, the character that varies most
dramatically among populations, as a potential isolating mechanism (although
this conclusion is based on the assumption that the populations are in
secondary contact). Therefore, that marcapatae
has a chestnut crown, in contrast to neighboring weskei and albiceps,
in itself provides no evidence for reproductive isolation.”
I
went on to conclude that changing currently recognized species limits was
unwarranted given the lack of critical data.
New
information:
Del Hoyo & Collar (2014) treated marcapatae
as a separate species based on the Tobias et al. point scheme as follows (provided
by Pam Rasmussen):
“Hitherto
treated as conspecific with C. marcapatae, but differs in its white
crown (3); stronger black lateral crownstripe (1); and much more evenly
delivered song, shorter in length (2), faster (at least 2), lower maximum
frequency (ns[2]), and different note shape (ns[at least 1]) (1). Monotypic.”
This
is a good example of the conceptual flaws of the Tobias et al. scheme, namely
that the scores are derived independent of phylogeny. In this case, we already know in sister taxa
that crown color is not associated with a barrier to gene flow, yet here it is
accorded 3 points out of the 7 needed for species rank.
The
vocal data are from Boesman (2016i), who analyzed 7
recordings of weskei and 9 of marcapatae:
“Song of weskei
differs from marcapatae
by much more uniform delivery, shorter length (score=2), lower max. frequency
(score=2), faster pace (score=2 or 3), descending song strophe (vs. rising and
falling, score 2) and different note shape (score 1). This would lead to a
total vocal score of 4 or 5 when applying Tobias criteria.”
On
the surface, these sound like impressive differences. I am assuming that the recordings represent
different individuals, but I leave it to those more experienced with Cranioleuca
voices to evaluate them.
As
for the inevitable demand for genetic data, I continue to point out their
limited utility for assigning taxon rank in cases like this, i.e. allopatric
taxa in a monophyletic group. We
already know what the results would be – the populations WILL differ,
especially given the obvious phenotypic differences. So, how different do they have to be to be
treated as species? Genetic data measure largely neutral loci not explicitly
those genes associated with gene flow and reproductive isolation, and thus
largely measure time-since-isolation and effective population size. The former is very roughly correlated with
speciation, but we have concrete examples of dramatic differences in genetic
distance among populations that are given different taxonomic ranks that no one
would dispute (e.g. capuchino Sporophila seedeater species vs. Syndactylus
bulbul populations that are phenotypically indistinguishable and not accorded
any taxonomic rank) that provide evidence that characters associated with
reproductive isolation do not evolve at a steady rate but instead
idiosyncratically. Looking at branch
lengths among close relatives eliminates some but not all of the problems. Anyway, see the SACC note on earlier genetic
studies that only show that we’re dealing with a monophyletic group, although weskei
to my knowledge has yet to be sampled.
Here
is the tree from Harvey et al. (2020), which shows clearly that albiceps
s.l. and marcapatae are sister taxa and that the node that unites them
is as deep as those linking most Cranioleuca treated as species, thus in
the crude comparative sense supporting species rank for them.

As
for evidence for or against gene flow between marcapatae and weskei,
these are strongly allopatric taxa. The
aerial distance on the map between the Vilcabamba range and the main Andes
might not be impressive to those not familiar with the area, but the ecological
gap is huge for high-elevation cloud-forest birds whose short, rounded wings
predict limited dispersal abilities.
Although in my opinion the dispersal ability of nonmigratory birds is
vastly underappreciated, nonetheless, I suspect that it would be so infrequent
between these two as to be impractical to detect without massive sampling.
Recommendation: Given my involvement
with these taxa, I think the right thing to do is recuse myself from voting,
and so I have no recommendation, and Glenn Seeholzer, whose dissertation was on
the Cranioleuca antisiensis group (e.g. Seeholzer & Brumfield 2018),
has agreed to take my voting slot. I
don’t have strong feelings either way at this point – I can see both sides.
English
names: If the proposal passes, then I see no need
for a separate proposal. This taxon has
no phenotypic features that distinguish it uniquely from other Cranioleuca,
and so Vilcabamba Spinetail seems not only the obvious choice but the only
one. Of course, if you see it otherwise,
please speak out and do a proposal.
References: (see SACC
Bibliography
for standard references)
BOESMAN, P.
2016i. Notes on the vocalizations
of Marcapata Spinetail (Cranioleuca
marcapatae). HBW Alive Ornithological Note 100. In: Handbook of the
Birds of the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100100
REMSEN, J. V., JR. 1984. Geographic variation, zoogeography, and
possible rapid evolution in some Cranioleuca spinetails (Furnariidae) of
the Andes. Wilson Bulletin 96:
515-523.
SEEHOLZER, AND
R. T. BRUMFIELD. 2018. Isolation by
distance, not incipient ecological speciation, explains genetic differentiation
in an Andean songbird (Aves: Furnariidae: Cranioleuca
antisiensis, Line-cheeked Spinetail) despite near threefold body size
change across an environmental gradient.
Molecular Ecology 27: 279-296.
VAURIE, C. 1980. Taxonomy and
geographical distribution of the Furnariidae (Aves, Passeriformes). Bulletin American Museum of Natural History
166: 1-357.
Van Remsen, June 2024
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Robbins:
“As far as new data, in my opinion, only the vocal data are potentially
relevant. Listening to vocalizations I think Boesman accurately describes
differences, but whether these mean anything who knows. I’m going to wait and
see what Glenn has to say on this given his deeper perspective on Cranioleuca
before I vote.”
Comments
from Jaramillo:
“NO for now, will wait until Glen Seeholzer gives and
opinion, I am open to change.”
Comments
from Lane:
“NO. This is a case that I think is a bit hobbled by
limited information. The distribution of the taxon weskei is still
largely thought to be solely in the Vilcabamba range of Cusco and Junin depts,
but since the 00's, it's been known that birds with white crowns occur all the
way to the north side of the Mantaro valley in Junin dept. This is the
population that I believe the recordings used in Boesman's assessment are from
<https://xeno-canto.org/species/Cranioleuca-marcapatae?query=ssp:%22weskei%22>. A
confounding factor is that there seems to be a vocal shift *within*
white-crowned birds across the Mantaro, as birds I recorded in 2010 in
Huancavelica dept (south of the Mantaro <https://macaulaylibrary.org/asset/392841541>) and
birds that Mark Robbins and Peter Hosner recorded in Ayacucho in 2012 <https://macaulaylibrary.org/asset/173820, https://macaulaylibrary.org/asset/173847> and in
the southern Vilcabamba in Cusco in 2012 <https://macaulaylibrary.org/asset/174006,https://macaulaylibrary.org/asset/186972> sound
considerably more like nominate marcapatae. We don't have recordings from the
type locality of C. m. weskei, so we can only assume that these more
southerly recordings are representative of the topotypical voice of the taxon.
So what does this apparent vocal shift mean across the Mantaro mean? Well, we
have no specimen material to review, but I suspect there may be an undescribed
taxon involved with the birds north of the valley, so Boesman's assessment of
voice doesn't actually apply to true C. m. weskei, but probably to
another taxon altogether which looks very like it. In the end, I think that
situation needs to be sorted out before we can make species-level changes in
the taxonomy of the C. marcapatae group. So, NO to splitting for now.”
Comments
from Glenn Seeholzer (voting for Remsen): “YES.
“At
the time of its description Cranioleuca
marcapatae weskei was only known
from the isolated northern Cordillera Vilcabamba. C. m. weskei and C. m. marcapatae
(hereafter weskei and marcapatae) display the same plumage
polarity (white vs. chestnut crown) that was used to diagnose the subspecies of
C. albiceps. The initial placement of
weskei as a subspecies of marcapatae was therefore based on the
comparative BSC approach - white vs. buffy crown coloration was not a
pre-mating barrier in C. albiceps
(widespread occurrence of intermediate crowns) therefore weskei and marcapatae
would also not exhibit reproductive isolation should they come into contact.
Specifically, the presence of extensive intermediate crown coloration between
the geographic and phenotypic extremes of C.
a. discolor and C. a. albiceps was considered a sign of gene flow. By this
same logic, if populations of weskei
and marcapatae come into contact and
show no evidence of intermediate crown coloration (i.e. introgression) this
would indicate that gene flow between the taxa is minimal and that weskei is a species.
“Variation in crown color in C. marcapatae
“Weskei was described based on
crown color – white – relative to the chestnut-crowned nominate subspecies
(Remsen 1984). Based on the specimens available at the time Remsen considered
crown color to be a discrete trait in C.
marcapatae in contrast to the continuous variation in crown color between
the buffy discolor extreme and the
white nominate subspecies of Cranioleuca
albiceps. Examination of recent specimens and ML photos (N=191) reaffirms
Van’s original assessment that there is no evidence of intermediate crown
coloration in either subspecies of C.
marcapatae, even where there come into closest contact in the southern
Cordillera Vilcabamba.
“See
the following ML image galleries
●
weskei (north
of the Río Montaro)
●
weskei (south
of the Río Montaro, west of the Río Apurímac)
●
weskei near
contact zone (east of the Río Apurímac)
●
marcapatae near
contact zone (west of -72°W)
●
marcapatae away
from contact zone (east of -72°W)
●
“The
ML photos fill in a large gap in specimen sampling in the southern Cordillera
Vilcambamba (Figure 1), and the two subspecies are now documented to occur
within ~40 linear kilometers of each other (60-70 km following the 3000 m
contour).
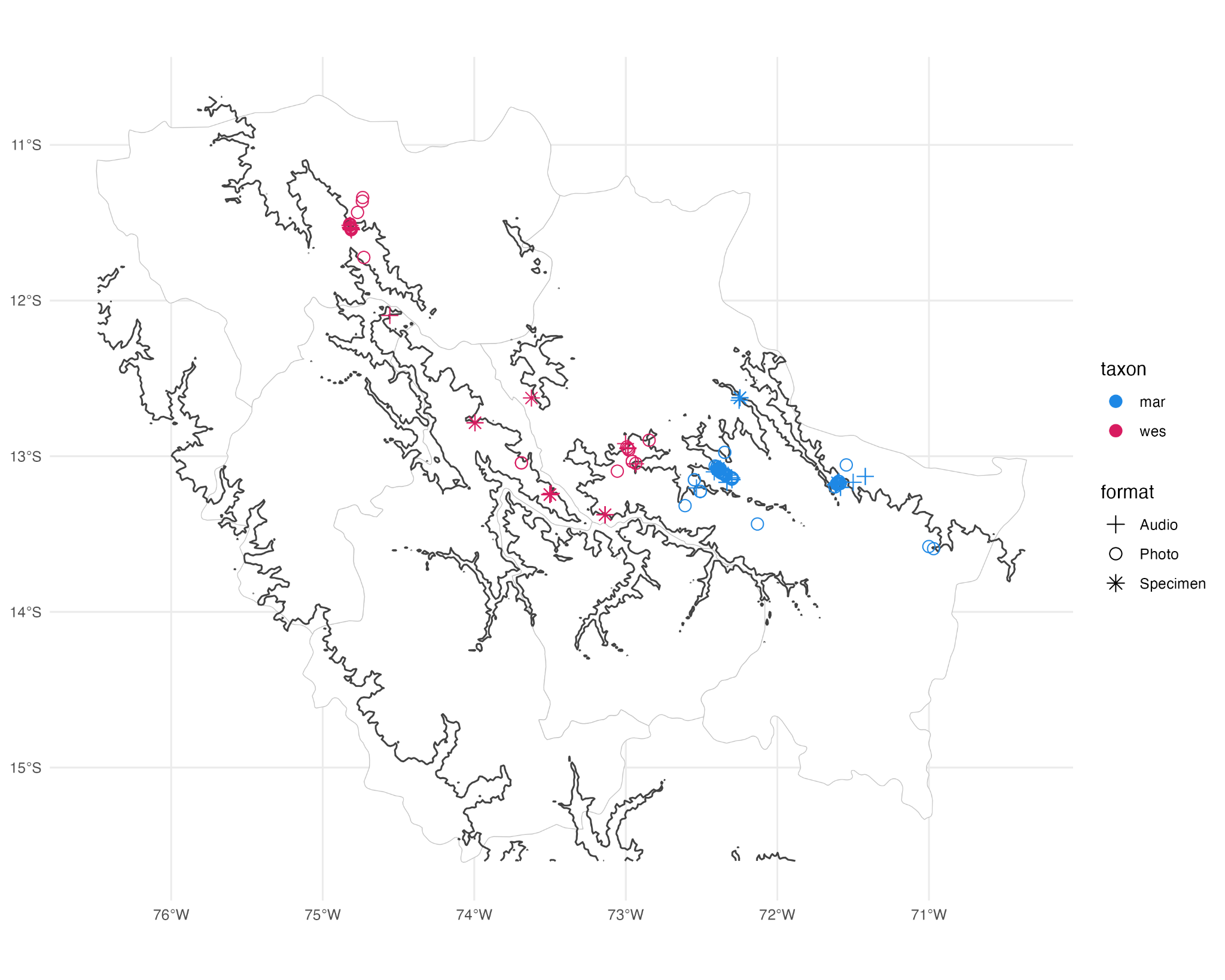
“Figure 1 - Map of vouchered records of Cranioleuca marcapatae. Color indicates
subspecies, shape indicates documentation method. Dark lines are 3000 m
elevational contours. Gray lines are departmental boundaries.
Below are the two closest photographs with
clear views of the crown.
“marcapatae
●
ML440773301 - Inca Trail above
Macchu Picchu (-13.226, -72.509)
●
ML217863001 - Camino Salcantay
above Colcapampa (-13.318, -72.609) - even closer than above but can’t confirm
that the crown is not intermediate
“weskei
●
ML492931841 - headwaters of the
Rio Vilcabamba near Lucma (-13.046, -72.934)
“These
localities occur at the headwaters of the Ríos Vilcabamba and Urubamba which
converge downstream of Machu Picchu. Examination of the satellite imagery of
the massif between these two rivers shows no obvious break in this species’
habitat (2500-3500 m montane forest). Unvouchered eBird observational records (S59715216, S33942238, S127451093) from near Mutuypampa
(-13.181,
-72.718)
occur midway along the 3000 m contour between the closest vouchered localities.
Given that weskei occurs across
significant habitat breaks elsewhere in its range (e.g. Río Apurimac, Río
Montaro), it’s a safe bet that some flavor of C. marcapatae continuously occupies the 2500-3500 m habitat band
between these rivers making this region a contact zone.
“For
comparison, specimens of Cranioleuca
albiceps from southern Puno to Cochabamba show the full spectrum of
intermediate crown coloration from white to buffy (see Van’s specimen series)
while spanning almost 500 linear km and numerous breaks in the 2500-3500 m
habitat band. Lest we doubt the ability of photographs to document intermediate
crown coloration, see this photo of two C. albiceps individuals next to each
other showing two different points on the white to buffy spectrum or this intermediate
individual.
“If
there were intermediates between weskei
and marcapatae outside of the
unsampled contact zone we would probably know it by now.
“Contact zone scenarios
“As
explained above, it is likely that weskei
and marcapatae come into contact
somewhere along the 2500-3500 m habitat band between the Ríos Vilcabamba and
Urubamba. There is a spectrum of possible patterns that we could expect in this
contact zone. At one extreme is assortative mating indicating reproductive isolation,
a slam dunk for treating weskei as a
species.
“At
the other extreme is a hybrid zone with panmixia creating a highly variable
population of individuals with intermediate crown coloration as seen in C. albiceps. If there is a hybrid zone,
it is a narrow one. From a strict BSC interpretation a hybrid zone with
panmixia demonstrates a lack of reproductive isolation and the interacting taxa
should be considered the same species. However, in practice we are highly
tolerant of this kind of hybridization between ‘good’ species.
“In
Cranioleuca, Seeholzer and Brumfield
(2017) documented a ~1000 km gradient in body size, plumage, and genetics in C. antisiensis which resulted in the
southern baroni and northern antisiensis groups being lumped. This
gradient was highly correlated with elevation and temperature and is best
considered a cline, not a hybrid zone. The only well-documented hybrid zone in Cranioleuca is between C. pyrrhophia and obsoleta. A thorough phenotypic study showed it to be 100s of
kilometers wide with extensive intermixing of pyrrhophia and obsoleta
phenotypic traits. Claramunt (2002) concluded that either the hybrid zone was
young or that some form of reproductive isolation exists maintaining the
phenotypes of the taxa in the face of gene flow. In either case, Claramunt
recommended the taxa continue to be treated as separate species.
“More
broadly we treat the members of many other super-species complexes with
well-documented hybrid zones as separate species. Manacus candei, vitellinus,
aurantiacus, and manacus all hybridize where their ranges abu and despite diverse
treatments we currently consider these all separate species. Other examples
include Rhegmatorhina berlepschi/hoffmansi, Myioborus melanocephalus/ornatus, Ramphocelus carbo/melanogaster, Thraupis
episcopus/sayaca, and of course most of the taxa that hybridize across the
Great Plains suture zone.
“I’m
too checked out of the speciation and taxonomy literature to parse the
meandering rationales for maintaining hybridizing taxa as species but if we’re
going to be consistent, then a narrow hybrid zone between weskei and marcapatae
would not be incompatible with treating them as species.
“Vocalizations
“After
listening to recordings, I agree with Dan that southern weskei and nominate marcapatae
sound very similar. Here are links to four galleries of ML recordings of the
short song (the most commonly recorded vocalization) so others can assess for
themselves.
●
weskei
(north of the Río Montaro)
●
weskei
(south of the Río Montaro, west of the Río Apurímac)
●
weskei
near contact zone (east of the Río Apurímac)
●
marcapatae near
contact zone (west of -72°W)
●
marcapatae away
from contact zone (east of -72°W)
“I
don’t have time to do a quantitative analysis, but I also don’t think it’s
necessary in this case. Absent an obvious diagnostic plumage feature like crown
color, the minimal vocal differentiation between southern weskei and nominate marcapatae
would indeed point us to one species. BUT we do have an obvious diagnostic
plumage feature that shows no evidence of introgression despite close contact.
A diagnostic vocal difference would help the case for species status, but the
absence of vocal differences doesn’t detract from it.
“Dan’s
intriguing observation that weskei
north of the Montaro in Junín sound different is not relevant to the question
of reproductive isolation or diagnostic characters between weskei and marcapatae
where their ranges abut. I will add that in addition to that population having
a higher, faster short song, the image galleries for this population also seem
to show a more extensive buffy wash to their throats compared to individuals
east of the Río Apurímac. Certainly seems like it could be an undescribed taxon.
“Summary
“Weskei and marcapatae show a discrete difference in
crown coloration (white vs. chestnut) with closely abutting distributions and a
high likelihood of a contact zone. This contact zone does not occur at an
ecotone or biogeographic barrier. Song divergence is minimal but not relevant
to assessing gene flow when crown color can be used as a proxy. There is no
evidence of introgression in crown coloration suggesting that in the contact
zone there is 1) assortative mating or 2) a narrow hybrid zone without gene
flow into either taxon. Clearly additional specimens and photographs from the
presumed contact zone are desirable but this would not change the patterns
outlined above which are enough for me to consider weskei a distinct species from marcapatae.
“Thoughts
“It
was fun to think about Cranioleuca
again after so many years. Specimens are always the most desirable form of
documentation. but I was impressed with how useful ML photographs were for this
system. For assessing the distribution of a simple binary trait like crown
color, they certainly came in handy!
“A
modern genomics treatment C.
marcapatae/weskei and C. albiceps
would make a great dissertation project. Replicate systems with what appear to
be two different evolutionary outcomes. For the future intrepid graduate
student inspired by this system, here are some juicy looking sampling
localities in the contact zone.
●
Above
Huayrani (-13.070,
-72.829)
●
Achirayoc
(-13.062,
-72.765)
and trails above (-13.119,
-72.771)
●
Mutuypampa
(-13.181,
-72.718)
●
Colcapampa
(-13.319,
-72.669)
“Additionally,
C. marcapatae and C. albiceps presumably come into contact
somewhere between Marcapata, Cusco, and the Sandia Valley, Puno along with
potential contact zones for many other Bolivian Yungas endemics and their
northern counterparts. High priority region for some adventure ornithology.”
Comments from
Areta: “" I vote NO to the
split. After seeing the case of C. baroni-antisiensis, taking a look at the
intergradation in crown color in C.
albicapilla (albicapilla
grades into albigula,
according to the HBW), knowing about the drastic geographic variation in
plumage and the dramatic within-locality vocal variation in C. pyrrhophia, thinking on
the geographic variation in crown color as exemplified by C. albiceps by Remsen, and
being absolutely convinced that the Boesman analysis is woefully incomplete for
a Cranioleuca I
don´t think there is a strong case here.
“Note:
there are sheer amounts on variation of speed of delivery, frequency patterns,
note shape, duration and pretty much any aspect one may want to consider even
in successive "songs" of the same individuals of Cranioleuca; therefore, any
analysis will need to a) have large sample sizes and b) take extra care to
compare the same vocalization types under the same motivational context [even
then, it might be mare]). So, I think that the vocal analysis needs to be
seriously downplayed here, given the high chances that it mischaracterizes the
differences and similarities in the voices of weskei and marcapatae.
Based on a cursory investigation of sounds, I don´t find any striking
difference in the "double-calls" or in the "short-song" of
these taxa. Of course, this is just a quick-and-dirty analysis, but I am
underwhelmed by the differences (unsurprisingly, given how difficult to
separate on vocalizations are several species of Cranioleuca...).
“Also,
there is this report by Hosner et al. (2015 WJO 127:563–581
, p. 575):
‘White-crowned'
Marcapata Spinetail (Cranioleuca marcapatae weskei).—We found this
spinetail uncommon to fairly common at all forested sites at 2,800–3,600 m (KU
112900, 112843, 122581–2 and CORBIDI LCA 2012-24, MCCF 476, MBR 8315, MBR 8339;
MJA 322, ML 173820, 173847, 174006–7, 174037, 186915, 186972). In June at Ccano,
a few non-vocalizing individuals were encountered. However, in September and
October at Chupón and above Paccaypata, presumed pairs and family groups (up to
4 individuals) were seen and heard daily as they foraged in Chusquea
bamboo understory at 3,000–3,600 m. The Ccano and Chupón records are the first
for Ayacucho. Cranioleuca m. weskei has also been documented recently
in Junín (e.g., XC 74478, 155740). Specimens from Paccaypata differed from
typical C. m. weskei in that the face and lores were plain gray,
rather than buffy, and a few white crown feathers had faint rufous edging.
These characters suggest that birds in Paccaypata may be intergrades between C.
m. weskei and nominate C. m. marcapatae from further east in
Cuzco. Apparent intergrades with a mix of rufous and white crown feathers have
been observed around Vitcos (13.111º S, 72.938º W, 3,000 m; Jul 2014; B.
Walker, pers. comm.)’. (Thanks to Tom Schulenberg for pointing this out in our
WGAC discussion).
“Thus, I see no reason to split weskei at present.”
Comments from Stiles: “NO on current
information. Given the variation in crown color within the close relative albiceps,
and with its white crown the primary basis for recognizing weskei as a
separate species, at least more genetic data and information on exactly where
recordings of several (and which?) taxa
were made will be needed to sort this out.”
Comments from Robbins: “After reading all the comments, including a number since I
first reviewed this proposal has me still on the fence. Nonetheless, if we must make a decision based
on current information I think it is premature to treat weskei as a species,
thus I vote NO for now.”
Comments
from Claramunt:
“YES. A difficult case as there are many in the genus Cranioleuca. But I would ask those that
voted “NO” to reconsider. Glen explained several key aspects very well. My
impression is that in the context of Cranioleuca taxonomy, weskei is clearly a different species. I totally
understand why Van described it as a subspecies: an isolated peripheral
population with a single (albeit conspicuous) plumage difference that happened
to be a variable trait in the sister species. Now we know that the distribution
of weskei is much larger, even larger than
that of marcapatae (still a minor point). But consider
the following:
1) It is not
uncommon for good species in Cranioleuca to differ in a single trait affecting the color
of the head: C. obsoleta
– C. pallida, C. erythrops – C. curtata. C. marcapatae and C. weskei would be just another case of that.
2) Crown color has great potential for
affecting mate choice and determining reproductive isolation in Cranioleuca and spinetails in general. As pointed out
before, different species of Cranioleuca typically differ more in plumage than in songs.
3) While albiceps shows a gradation in the color of the crown,
the crown varies from white to light cinnamon only, not to rufous like the
crown of marcapatae. The
magnitude of the difference is much greater in the case of marcapatae and weskei.
4) Also in contrast to the albiceps case, the difference between marcapatae and weskei is discrete; intermediate phenotypes have not
been described so far, with the exception of the mention of specimens of weskei from one locality in which “… a few white crown
feathers had faint rufous edging,” and a report of another population with
birds with crowns with mixed crowns (Hosner et al. 2015). I would take those
observations with a grain of salt until more details are published.
5) As pointed out by Glenn, if there
is indeed some hybridization, it seems very limited geographically.
“Future work on these species will tell what is exactly going on,
but in the context of our current taxonomy of Cranioleuca, I think that weskei should be treated as a species.”
Additional
comments from Stiles:
“I find Santiago's comments very interesting,
especially his overview of the main part of the distribution of Cranioleuca
- hence, I am willing to change my vote to YES for proposal 1010.
Comments
from Ryan Terrill (voting for Del Rio): “YES -- I think this is a very borderline case
that warrants more data and could go either way, but Glenn's comments help a
lot here and are appreciated. It does seem like there is likely to be a contact
zone somewhere in the headwaters of the Vilcabamba/Urubamba rivers, which would
be great to visit and sample. But, as noted, if they do come into contact, they
either don't interbreed, or if they do, form a very narrow hybrid zone. Other
examples of variation within Cranioleuca species show a lot of variation
and trait introgression – notably the C. antisiensis and C. albiceps
complexes. In reference to
Santiago's comments about crown color as a trait affecting mate choice, I have
seen pairs of C. albiceps segregate by crown color within their contact
zone in Cochabamba, see my notes in this checklist -- https://ebird.org/checklist/S42501143 "A
pair of birds near the top of the road had pale yellow crowns, noticeably and
distinctively paler than the orange crowned birds just down the road". Of course, this is
only one anecdote.”
Comments
from Bonaccorso:
“YES, with hesitation, but Santiago’s and Glenn’s arguments are compelling. It
is unfortunate that more genetic data from additional individuals and formally
analyzed songs are not yet available.
“Regarding
the crown variation in albiceps, I don’t consider it a problem. It’s
plausible that this variation existed in ancestral weskei but was lost
with the fixation of the white crown. This is consistent with an isolated
population in a small range, where drift and sexual selection could drive rapid
evolution. I also agree with Santiago that a white crown could play a crucial
role in pre-zygotic isolation. Combined with the long genetic branch between weskei
and marcapatae, these points support a split.”
Comments
from Zimmer:
“YES. As
others have noted, this one is something of a borderline case in terms of
supporting evidence, one way or the other.
I listened to all of the linked vocal samples that Glenn provided, and I
remain unimpressed with any alleged vocal differences between weskei and
marcapatae. That having been
said, I could say the same about the songs of many species-pairs within Cranioleuca -- so many of them are extremely similar in
songs, and I’ve noted marked variation from one individual to the next from
within the same population, and even from one song to the next from the same
individual, particularly depending on state of arousal (e.g. in response to
playback). Throw in the fact that many
species in the genus duet, and, that you’ve got no sexual dimorphism to
distinguish the sex of a lone vocalizing bird, and it means that even a
detailed quantitative vocal analysis will have a hard time controlling for sex
and number of individuals when analyzing archived recordings from third parties. So, I would agree with Glenn that the absence
of a quantitative vocal analysis in this case is not a deal-breaker, given that
we do have an obvious, discrete and diagnostic plumage feature, for which we
have no evidence of introgression despite close contact. To this end, I am persuaded both by Glenn’s
reasoning, and, particularly, by the points made by Santiago regarding the
potential for crown color or other single plumage characters affecting mate
choice in Cranioleuca species-pairs (and the examples Santiago offered
are spot-on in my opinion – my mind went immediately to pallida versus obsolete,
whose songs I can never personally discriminate in the field), and in the
discrete nature of the rufous versus white crown color in marcapatae
versus weskei, as opposed to the continuous variation in albiceps. Also, as pointed out by Glenn, Ryan and
Santiago, assuming there is a contact zone between weskei and marcapatae,
the two are either not interbreeding at all, or, the hybrid zone, if it exists,
must be extremely narrow, given the complete lack of evidence for phenotypic
introgression in crown color.”
Additional
comments from Jaramillo: “I just did a quick read through of the proposal, and the
information I had not seen before. I am switching to a YES on Cranioleuca
weskei based on Glenn's information and Santiago's analysis of Cranioleuca
in general. It may shift in the future as we obtain more information, but I am
comfortable with a YES vote based on what I read and understand right now.”