Proposal (1012) to South
American Classification Committee
Treat Sphenopsis
ochracea and Sphenopsis piurae as separate species from S.
melanotis
Note: This is a
high-priority issue for WGAC.
Background: Our SACC note on this is as follows:
10. Despite strong differences in some plumage
characters, Sphenopsis (formerly Hemispingus) melanotis
has been treated a polytypic species based on shared plumage themes (Hellmayr
1936, Zimmer 1947, Storer 1970a, Meyer de Schauensee 1970, Dickinson &
Christidis 2014. and others).
García-Moreno et al. (2001) and García-Moreno & Fjeldså (2003) found
that the distinctive taxon piurae, currently treated as a subspecies of Sphenopsis
melanotis (e.g., Meyer de Schauensee 1970), is more distant from the latter
than is S. frontalis, and that piurae is basal to frontalis +
melanotis; these analyses, however, were based on only ca. 300 base-pairs
of mtDNA. Ridgely & Greenfield
(2001) treated piurae as a separate species from H. melanotis
based on plumage and vocal differences. SACC proposal to
recognize piurae as a species did not pass. Hilty (2011) also treated piurae as a
separate species. Ridgely &
Greenfield (2001) and Hilty (2011) further recognized the subspecies ochracea
as a separate species based on plumage differences. Halley (2022) treated piurae and ochracea
as species based on distinctiveness of plumage.
SACC proposal needed.
We
currently treat Sphenopsis (ex-Hemispingus) melanotis
(Black-eared Hemispingus) as a highly polytypic species, as did Dickinson &
Christidis (2014), who recognized 6 subspecies in 3 groups in the humid Andes:
(1) nominate melanotis
(Andes of NW Venezuela to e. Ecuador), berlepschi (e. Peru), and castaneicollis
(Eastern Andes of S. Peru and Bolivia)
(2) ochracea (
Western Andes of sw. Colombia and nw. Ecuador)
(3) piurae (s.
Ecuador, NW Peru) and macrophrys (Western Andes of c. Peru).
We
rejected a proposal in 2007 (SACC 284) based largely on weak
genetic data and absence of data on vocalizations.
Here
is a crude home-made representation of the distribution of the taxa in
question. All photos are from Macaulay
Library (melanotis by Ben Jesup, ochracea by Angel Argüello
Méndez, piurae and macrophrys by Fernando Angulo, berlepschi
by José Martín, castaneicollis by Tini & Jacob Wijpkema). Note the subspecies macrophrys Koepcke 1961 has been generally
overlooked (“Near piurae but with broader white
superciliaries, a broader and more conspicuous gray band on nape and
post-auriculars, and underside of wing more whitish”). See also Halley (2022) for specimen photos.
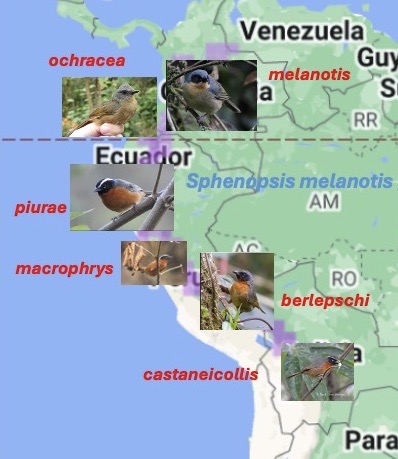
From
the differences in plumage, one can quickly spot the problems. It is not immediately obvious why ochracea
was ever considered conspecific, although the rationale was likely that it
looks like a typical smudgy, obscure Chocó representative of a group of more
brightly colored taxa. It also could be
argued that nominate melanotis stands apart from all the rest. From the perspective of those most familiar with
birds north of the Marañon, such as Ridgely and Hilty (see SACC Note), one
might immediately reject piurae as being conspecific with nearby
nominate melanotis because of the plumage differences, but for those
more familiar with taxa south of the Marañon, such as J. Zimmer and Schulenberg
et al. (“Birds of Peru”), piurae looks similar to distant S. m.
castaneicollis. I suspect that a
formal analysis of plumage similarities could go any number of ways depending
on how one scores the plumage characters, e.g. see Halley (2022: 221). Lots of “eye of beholder” reasoning would be
involved. It seems we have a classic
conundrum concerning taxon rank of distinctive allotaxa, with the only clear
case being macrophrys as a subspecies regardless of species limits.
New
information:
Del Hoyo & Collar (2014) treated ochracea
and piurae as a separate species based on the Tobias et al. point
scheme as follows (provided by Pam Rasmussen):
“Western:
https://birdsoftheworld.org/bow/historic/hbw/bkehem3/1.0/introduction
[ochracea] Usually treated as conspecific with S. melanotis
and S. piurae, but each differs markedly in plumage and habitat and
hence is tentatively accorded species rank here (although songs of present
species and melanotis difficult to distinguish (1) ); review of group
warranted. Monotypic.
“Piura:
https://birdsoftheworld.org/bow/historic/hbw/bkehem1/1.0/introduction
Usually
treated as conspecific with S. melanotis and S. ochracea (see
latter); present species exhibits some vocal differences, and limited molecular
data suggest that it is at least as distinct from S. melanotis as latter
is from S. frontalis; review of group warranted. Two subspecies
recognized.”
The
habitat difference alluded to likely comes from Ridgely & Greenfield’s
statement: “unlike.
[melanotis and piurae], the Western Hemispingus does not show
any particular predilection for an understory of Chusquea bamboo.” However, photos of ochracea in
Macaulay show bamboo in 11 of the small number of photos of the species, so
perhaps that needs re-evaluation.
Certainly it should not play a role in determining species limits.
Boesman (2016k) analyzed an
unspecified number of recordings from unspecified locations. Apparently, vocalizations of berlepschi
and hanieli were not analyzed. Here
are his main conclusions:
“From the above, it is clear that duet song
of all three species is structurally similar. There seems to be however a
closer resemblance between duets of H. ochraceus and H. melanotis.”
“All in all, we can conclude that voice of H. ochraceus is about identical to H. melanotis.”
“Voice of piurae
at the other hand is quite distinctive
and can be safely told apart.”
“Race castaneicollis of H. melanotis
also differs from other races of this species”
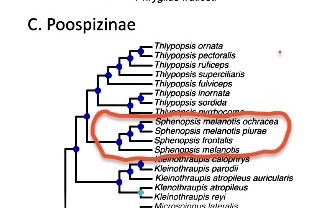
Price-Waldman
(2019), an unpublished dissertation from Kevin Burns’ lab, used UCE data to
construct a phylogeny of the tanagers.
As cited by Halley (2022), the topology for this group was (((ochracea,
piurae), frontalis), melanotis), which would thus require
separation, minimally, of ochracea + piurae as a separate species
from melanotis.
Expanding
frontalis to include all of the melanotis group is clearly
untenable because frontalis is broadly sympatric with melanotis
representatives. Tangentially, note that
the subspecies of frontalis of the Coastal Range of Venezuela, S. m.
hanieli, superficially looks as much like a melanotis type as a
representative of frontalis, and Halley (2022) emphasized the
distinctiveness of this taxon and its tentative placement within S.
frontalis. Here’s a photo from
Macaulay by Margaret Wieser from Aragua:

So,
species limits need to be changed, but what do we do with berlepschi, castaneicollis,
and even hanieli? Certainly by
extrapolation, the differences in plumage between nominate melanotis and
berlepschi + castaneicollis are of the same general magnitude as
those between nominate melanotis and ochracea, again depending on
how one weights characters and determines homoplasy.
Halley
(2022) made two important contributions.
First, he pointed out that the Garcia-Moreno et al. data-set actually
lacked a sample of ochracea despite its claims otherwise, and second, he
assembled the few existing specimens of ochracea to show that it had
been inaccurately illustrated in all published works, with respect to
coloration of the underparts, which are indeed ochraceous, and in some cases
with respect to having a mask, which is minimal. Halley did some solid, baseline
alpha-taxonomy that will be useful for all future analyses.
Discussion
and Recommendation:
Just to understand the history of all this, I have recorded here my wanderings
down several rabbit holes (to use a worn-out metaphor). I was tempted just to delete everything and
just present the Price-Waldman tree, which requires at least a 2-way
split. But that makes me
uncomfortable. The thesis is
unpublished. Given the surprising
finding, were vouchers double-checked?
And what about unsampled berlepschi-castaneicollis, for
which Boesman has evidence for vocal distinctiveness (at least for castaneicollis)
and which, as a group, are phenotypically distinctive? Do we keep them with melanotis despite
plumage and vocal differences? And what about the largely neglected hanieli
issue? My gut instinct is not to meddle
with our current classification, despite the possibility of a paraphyletic melanotis,
until all these other issues are sorted out.
In contrast to the philosophy of many colleagues, I actually like
conflict among world classifications because it signals honestly and appropriately
that there is considerable uncertainty, which would otherwise be masked. Nevertheless, I don’t have a strong
recommendation and will wait to see what others say.
English
names: “Western Hemispingus” and “Piura Hemispingus”
have more than two decades of traction in the literature. I suspect “Western” is not a name anyone
really likes. Hellmayr (1936) called it
“Ochraceous-bellied Hemispingus”. The
reason why Ridgely & Greenfield (2001) didn’t go with this is because they
described the underparts as “drab buffy olivaceous”, although Greenfield’s
illustration of the head shows the throat at least to be ochraceus. The ochraceous belly (vs. all of the
underparts) highlights what seems to be a unique feature (and it also helps
remember the scientific name ochracea).
I predict if Bob Ridgely knew what we do now about ochracea, he
would gave stuck with Ochraceous-bellied.
I am willing to write a short proposal on this if anyone else is favors
overturning 20+ years of stability. If
it were 50+ years of stability or if all classifications treated it as a
sperate species “Western Hemispingus”, then I would object to changing the
name, but if we are ever going to do it, now is the time (if the proposal
passes). “Ochraceous-bellied
Hemispingus” would have the additional advantage of pointing out the problems
with illustrations of ochracea, as noted by Halley (2022). In contrast, “Piura Hemispingus” was used by
Hellmayr (1936) and thus has been in the literature for at least ca. 88 years.
This dodges the problem that none of
these species are in the genus Hemispingus any longer, but that’s a
separate problem that would have to consider the broader diaspora of former Hemispingus. For now, I’m personally content with
“Hemispingus” as a vague morphotype. i.e. roughly as a “half finch”, as in the
derivation of the genus name, in the interests of stability, but if anyone
wants to tackle this problem, feel free.
References: (see SACC
Bibliography
for standard references)
HALLEY, M. R.
2022. Taxonomic status of the
Western Hemispingus Sphenopsis ochracea (Thraupidae) and a review of
species limits in the genus Sphenopsis P. L. Sclater, 1861. Bulletin British Ornithologists’ Club 142:
209-223.
ZIMMER, J. T. 1947. Studies of
Peruvian birds, No. 52. The genera Sericossypha, Chlorospingus,
Cnemoscopus, Hemispingus, Conothraupis, Chlorornis, Lamprospiza, Cissopis,
and Schistochlamys. American
Museum Novitates 1367: 1-26.
Price-Waldman, R. M. 2019. Phylogenomics, trait evolution, and
diversification of the tanagers (Aves:Thraupidae). M.Sc. thesis. San Diego State Univ.
Van Remsen, June 2024
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Robbins:
“NO. Another messy situation for all the reasons that are pointed out in the
proposal. Given the number of issues that need to be addressed, as summarized
by Van, for now I vote NO.”
Comments
from Jaramillo:
“YES – You only need to look at the map, compare to other similar Andean chain
“superspecies,” look at the photos and you have the informed suspicion that
more than one species is involved in melanotis. The unpublished (thus far) DNA work, the
vocal information that is available adds more pieces to the puzzle, and they
also lean in the way of separating this chain of taxa into multiple species. I
am comfortable in voting yes, and comfortable in understanding that in the
future there may be future changes to this complex.”
Comments from Areta: “A conflicted NO. I think that
there are at least 3 species in S.
melanotis (and probably more), but I feel quite uncomfortable with the lack
of data on berlepschi
and melanotis
(taxa sampled by Price-Waldman 2019: Sphenopsis
melanotis castaneicollis FMNH 430079 / Sphenopsis
melanotis ochracea ANSP 149722 / Sphenopsis
melanotis piurae FMNH 480966) and I don´t like to guess. berlepschi is clearly more
similar to melanotis,
so it might not be much of a problem (perhaps...), and I could accept that it
is conspecific with melanotis.
But castaneicollis,
with its broad black mask and chin, contrasting white supercilium and
rufous-chestnut large pectoral band is much more like the widely allopatric piurae, while also being
quite distinct from melanotis/berlepschi. Ochracea is ridiculously
different from the rest, yet based on Price-Waldman it is sister (how deeply
diverged? we don´t know) to the distinctive piurae,
both from the W slope of the Andes. Therefore, the drabbest and the most
colourful forms are sister, suggesting that concluding on the phylogenetic
placement of other taxa based on plumage is risky. Should we go with geography
then? Risky again. Then there are also Van´s worries about macrophrys and S. frontalis.
“This is one of the situations in which there is
(barely) enough information to split some of the taxa, but in which information
on key taxa is missing to properly establish species limits and in which we
must guess in order to decide. Shall we stick to the old taxonomy or move to a
newer one that may have other problems? Also, the Price-Waldman phylogeny has
not been adequately published, and all we seem to have at hand is the topology
of a tree. Halley (2022) provides an excellent overview of the taxa involved
and clarifies some problems while proposing an alternative taxonomy. Although
it seems clear that the single-species treatment will fall, I am not convinced
about how many species we should recognize and how each of them should be
confirmed. Therefore, I vote NO to any split for the time being.”
Comments
from Stiles:
“NO. The whole situation is simply at the stage where too little is known about
the genetics and vocalizations of too many taxa, and their geographic
distributions appear to present a more complicated picture. A more
comprehensive study of Sphenopsis is badly needed - splitting these two
now might be jumping the gun.”
Comments
from Lane:
“NO. Another situation where I think we are voting without all the necessary
information in hand to make an informed decision. What harm is there in waiting
for someone to compile those data?”
Comments
from Niels Krabbe (voting for Del-Rio): “NO. There is no indication that
Price-Waldman (2019) double-checked the identity of his frontalis (MSB
31856 (Peru: dpto. Amazonas; prov. Utcubamba; dist. Lonya Grande; ca. 4.5 km N
Tullanya). Halley (2022) certainly did not. However, as both Price-Waldman and
García-Moreno & Fjeldså (2012), the latter using a double-checked frontalis
from Imbabura, W Ecuador, found frontalis to be embedded in the melanotis
complex, there is no reason to doubt the correct identification of the Amazonas
specimen.
“A
recent, as yet unpublished find by Jonas Nilsson (pers. comm.) of a population
in the Chilla Mts of southern Ecuador (a ridge connecting the east slope with
the west slope) that appears intermediate in plumage between the vocally
similar ochracea and melanotis further underscores the need for a
detailed study that includes all the Sphenopsis forms before changing
the current classification.”
Comments from Claramunt: “YES. The only problem here is
our regressive tradition in ornithology of stuffing taxa into polytypic species
based on pure speculations about degree of differentiation. The actual
information we have is: 1) subspecies now included in melanotis are not each other’s close relatives because of
the position of S.
frontalis (Garcia-Moreno
et al, Price-Waldman). 2) Examination of study skins point to the existence of
three well delineated taxa (100% diagnosable, Halley 2022): melanotis, ochracea, and piurae. The solution is simple: split the
polytypic entity into its fundamental units. And the result is totally coherent
from a biogeographical point of view. I fail to see problems with the proposed
taxonomy. In contrast, I don’t see any evidence supporting the traditional
treatment. Where is the evidence that ochracea or piurae are reproductively compatible with melanotis?”
Comments
from Remsen:
“NO. I appreciate Santiago’s points above except I’m not sure what the
appropriate species-level components are.
As for evidence for reproductive compatibility among various taxa, we
could say that about every single case of allotaxa ranked as species for which
vocalizations have not been properly analyzed.
I can see how the Peters-era people could consider them conspecific
because they all share plumage features to varying degrees and are clearly
allopatric replacements. What they were
basing their decision on implicitly is that one can line up specimens of the
taxa from the dull extreme of ochracea to the bright extreme of piurae,
and see a series of taxa that bridge the extremes. The problem is that they don’t line up in a
strict geographic sequence.
My NO is based on the quality and
quantity of data available. We’ve got an
unpublished dissertation for which a voucher needs to be checked. We’ve got a
vocal analysis that did not report N or localities, with a critical missing
taxon. We’ve got the conceptually flawed
Tobias et al. scheme that may have used faulty information on habitat. The genetic analyses have missing taxa. The argument for multiple species is built on
a flimsy assembly of substandard data to the point that I think there is more
to be gained by retaining the traditional classification until more solid data
are produced than by endorsing one or more splits on the basis of weak data. On the other hand, I suspect that there are
multiple species-level taxa in this complex awaiting to be properly elucidated
once more rigorous data are produced.”
Comments
from Bonaccorso:
“NO, but hesitantly. As Santiago points out, the three units (melanotis,
ochracea, and piurae) are indeed diagnosable. However, we lack
sufficient knowledge about what might be occurring in a potential ochracea-piurae
contact zone in southern Ecuador (see Niels’ comments on Jonas Nilsson’s
unpublished data). This is a highly complex and understudied region that could
reveal fascinating patterns. We might be facing another conundrum like the Myioborus
ornatus-melanocephalus situation.”
Comments
from Zimmer:
“NO. I agree
with the thrust of Santiago’s comments overall, and feel pretty certain that
there are, indeed, multiple species-level taxa currently nested within this
single polytypic “species”. But, as Van
points out, there are critical flaws in all of the various analyses supporting
the proposed splits, and Niels’ comments regarding as yet unpublished
observations by Jonas Nilsson of a population in southern Ecuador that is
intermediate in plumage between ochracea and melanotis further
suggests that we should tap the breaks on making any premature moves until we
have more expansive data sets for all of the populations.”