Proposal (1018) to South
American Classification Committee
Treat Xenodacnis
petersi as a separate species from X. parina
Note: This is a
high-priority issue for WGAC.
Background: Our SACC note on this is as follows:
51a. The northern petersi subspecies group was formerly (e.g.,
Bond & Meyer de Schauensee 1939, Zimmer 1942) considered a separate species
from Xenodacnis parina. All
subsequent classifications since Meyer de Schauensee (1970) and Storer (1970a)
have treated them as conspecific. Del
Hoyo and Collar (2016) treated it as a separate species, with support from
vocal differences found by Boesman (2016m).
Aguilar (2021) found that the Ecuadorean population of X. p. bella
was significantly larger in body size and had a different habitat preference
(Aguilar 2019) than all other taxa.
To
set the stage for this one, here is the map from Aguilar (2021):
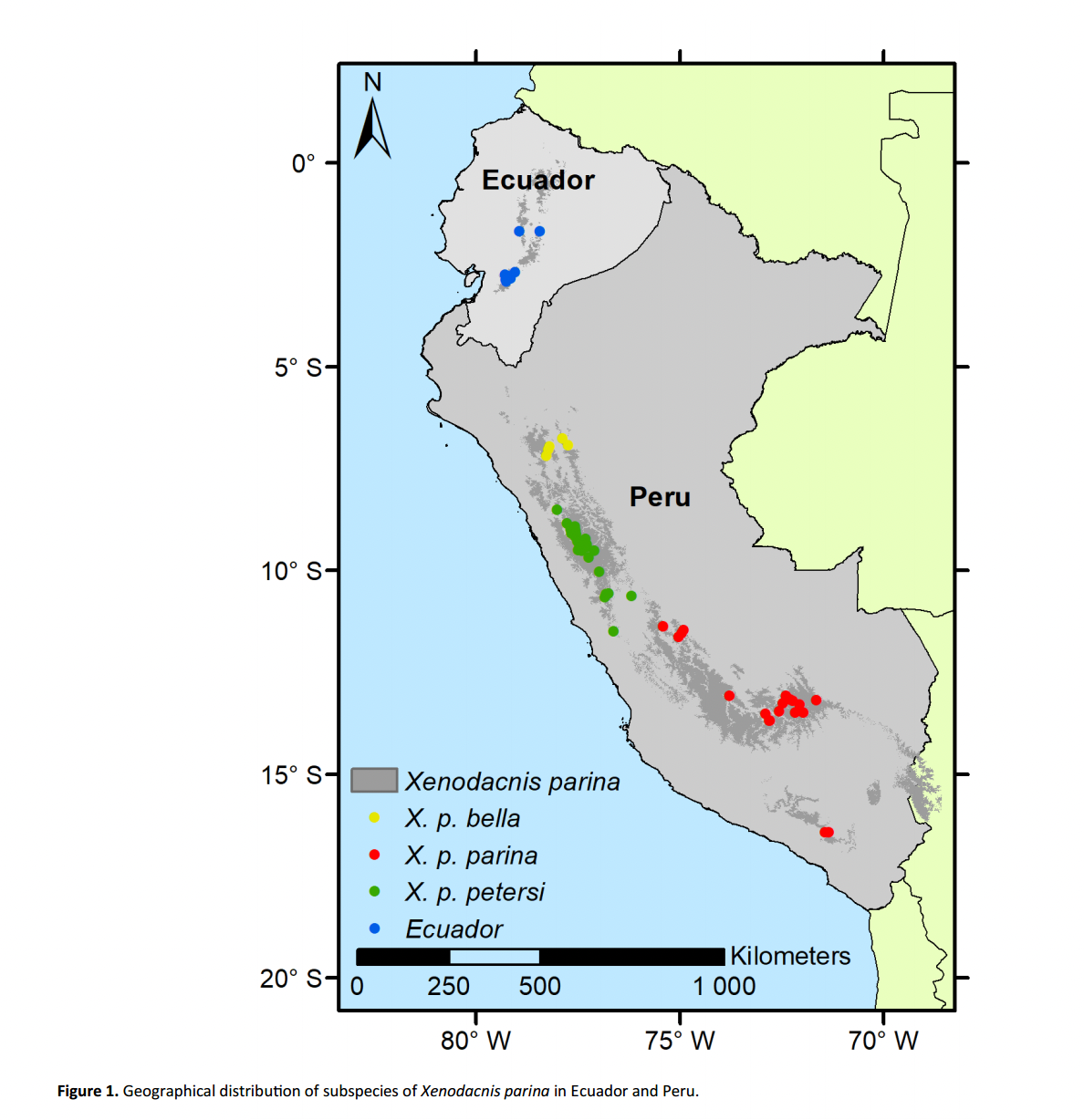
Long
treated as a monotypic species (e. g. Hellmayr 1935), a paucity of specimens of
this patchily distributed Polylepis-zone specialist likely prevented earlier
authors from noting the fairly pronounced geographic variation that was noted
by Bond & Meyer de Schauensee (1939), who described petersi as a
separate species from X. parina, and in the same publication
described bella as a subspecies of petersi. Zimmer (1942) also treated
petersi as a separate species. Zimmer
only had 1 petersi to examine but he noted:
“The
differences from X. parina are not very pronounced, except for the
considerably greater size of petersi. The bright streaks on a large part
of the blue feathering are suggested in parina though inconspicuously
so. The distinctions between the two forms are such that intergradation could
easily exist somewhere in the broad area intervening between the respective
ranges, but it has yet to be demonstrated. Until some closer approach in range
and taxonomic characters can be demonstrated, the two groups are best left
specifically distinct.”
Concerning
bella, Zimmer wrote:
“Not in
the collection before me. In a sense this form is intermediate between typical petersi
and parina, being smaller than petersi and with somewhat less
strongly marked bright streaks. The more greenish tinge of the male plumage
ought to be substantiated by more material since it is within the possible
bounds of individual (and "ageal")
variation, as demonstrated by the specimens of parina discussed
earlier.”
Here
are photos of LSUMZ specimens of the three taxa, males and females. I should have checked petersi vs. bella
carefully when I took the photos to see if the plumage difference I see in the
photos holds up, especially because they conflict with some statements
herein. There is a real size difference,
obvious to the eye, evident in both sexes.
(Many of you will recognize some of the preparators on the labels, all
officially Useful Humans. John O’Neill,
and even Mark Robbins, was skinning birds before some of you were born. Note also the cleverness with which I have
deftly offset petersi in the photos slightly to the left to highlight
that it is primarily a bird of the Western cordillera):



Bond
(1955) maintained the two as separate species.
(He also provided a tidbit on one of M. A. Carriker’s
[see previous proposal] above-average field days “It is of interest that
Mr. Carriker obtained a specimen of this interesting bird on the same day
[March 26, 1932] that he collected his types of Yanacea alpina and Leptasthenura
yanacensis and an example of the recently described Zaratornis
stresemanni Koepcke. His field notes for that day state that he ‘went up to
Puno’, and ‘worked up to about 15500 feet, but most of the shooting was done
between 14000 and 15000 feet.’")
Meyer
de Schauensee (1966) was the one who first lump then. Shaun Peters (in litt.) graciously dug out
this quote from that work (of which I no longer have a copy). From Meyer de
Schauensee (1966) under Xenodacnis parina:
“Includes
Xenodacnis petersi (Bond & de Schauensee, Not.
Naturae, 1939, no. 40, p.1, Yanac, Ancash, Peru) and
X. petersi bella (Bond & de Schauensee,
l.c., p.2, Atuen, Amazonas, Peru)
“Recently, W. G. George collected
specimens of this rare bird in southwestern Arequipa, Peru and found that birds
from this region combined characters of both
X. parina and X. petersi, showing
them to be conspecific.
“George thinks that this aberrant
honeycreeper is probably related to Dacnis. The pale
glistening blue stripes on the feathers of the back and chest are found in both
Dacnis berlepschi and
X. parina."
This
seems to have become the accepted subsequent classification (e.g. Storer
1970a). Now I really want to see those
George specimens at AMNH to assess the “combined characters” assessment. Shaun Peters again came to the rescue in
providing a jpg copy of George (1964), using Archives.org, and there is nothing
I can find in that article relevant to species limits.
Isler
& Isler (1987, The Tanagers) described the vocalizations in detail, but
this was based on Ted Parker recordings, which at that time might have been
based on petersi only; no differences among taxa were mentioned.
Fjeldså
& Krabbe (1990; Birds of the High Andes) wrote: “Vocalizations of ssp
parina considerably weaker than in other sspp.”,
although they considered bella “doubtfully distinct” from petersi.
Here
is the helpful account from Schulenberg et al. (2007; Birds of Peru guide) with
Dale Dyer’s plate:

New
information:
Del
Hoyo & Collar (2014) treated as a separate species based on the Tobias et
al. point scheme as follows (provided by Pam Rasmussen):
"Streaked:
https://birdsoftheworld.org/bow/historic/hbw/tildac3/1.0/introduction
Hitherto
treated as conspecific with X. parina, although given species rank when
first described, with good reason: differs in its (in male) bright blue shaft
streaks (present but less obvious on race bella) vs none (1); (in
female) mid-crown to nape ashy brown vs dull blue (3); blue vs ferruginous on
malar area at base of lower mandible (ns[1]); blue vs whitish subocular line
(1); much larger size (five male wings comprising nominate 82.5, 79, 78 mm and bella
76.5, 75.5 vs two X. parina 66.5, 61.5 (1); allow 3); moreover,
commonest song type very different (lower speed and lower number of notes:
2+2=4) (2). Race bella marginally smaller and duller. Names petersi
and bella described simultaneously, but latter as a race of former, so petersi
claims priority. Two subspecies recognized (but see below)."
The vocal data are from
Boesman (2016m), which presents
sonograms from what I assume are three different individuals each from the
Ecuador population, petersi, bella, and nominate parina,
plus one from the somewhat anomalous Arequipa population:
“Song of petersi and bella is a sequence of some 5-10 clear
whistles, typically one or more series of repeated identical notes, which are
either sharply upslurred or downslurred. Most notes reach max. frequencies of
8-10kHz.
“All but one recording of nominate are
very different from northern races, song having mainly peculiar subdued
lower-pitched bubbling notes at a much higher trilled pace. This would seem a
major vocal difference, but then there is a single recording from Arequipa
(ML33844 from Ted Parker), which is about identical to examples from Ecuador
(!). This greatly reduces the marked vocal difference, but nevertheless, it
would seem that the most commonly uttered song-type of nominate is very
different from any vocalization known from northern races. Based on high pace
and high number of notes, a 2+2 score could be given (albeit with some
reservation, as there may be two song-types involved).”

Note
that Boesman was placing the Arequipa population with nominate parina
based on its southerly distribution, but photos by Tom Schulenberg of George’s
specimens at AMNH clearly indicate that it belongs with petersi, thus
matching the vocal diagnosis.
Discussion
and Recommendation:
Here we have the Classic Allotaxa Conundrum, with only fragments of information
and data with which to work. There are
no contact zones to guide us – in fact, even within these taxa, the tiny
populations are all isolated from each other in habitat islands at high
elevations. Genetic data would be
fascinating, particularly since the effective population sizes are likely tiny.
Reasons
to vote YES: (1) No rationale has ever been published for the lump, or at least
I can’t find it; so, we would be returning to the original classification. (2)
Vocal differences as described by Boesman seem substantial. (3) The plumage
differences are substantial, and at the risk of getting touchy-feely, they feel
like different species to the touch, to me anyway; it’s not just color but
something also to do with texture, perhaps an artifact of the shaft streaking
in petersi/bella. Even the
females seem to have a different plumage quality.
Reasons
to vote NO: Are Boesman’s data sufficient for a split? He sampled all the populations, but the total
N is unknown, and I would certainly feel more comfortable with a formal
analysis that also specified no doubt on homology of vocalizations. How soft are we going to get in terms of
requiring a formal analysis in a potentially complicated issue like this
one? I hope someone here has the energy
to explore recordings on xeno-canto and Macaulay, which are likely many more
now than were available to Boesman. By
the way, for those of you not on eBird, you can do this efficiently by going to
eBird explore page, then hit “Search
Photos and Sounds” in lower right; note that you may have to hit refresh to get
anything to appear at all; then, enter a taxon name
and under “More filters” check “song” or “call”, and where it says Recently
Uploaded on far right, toggle that to Best Quality (although beware that eBird
recordists, like eBird photographers, chronically over-rate the quality; also
that not all recordings are rated, so it might also be worth toggling to Least
Rated and skimming through sonograms for good ones).
I
have no strong recommendation, although I lean slightly towards a YES. I have conscripted Niels Krabbe to take my
vote on this one.
English
names:
BirdLife International used “Tit-like Dacnis” for nominate parina, and
“Streaked Dacnis” for the petersi group.
I have problems with this. If
there was only one species, then Tit-like Dacnis works pretty well. The “Tit-like” combined with the genus name Xenodacnis
(“foreign Dacnis”, with xeno being one of the Greek roots that is more
familiar in meaning through “xenophobia”) adequately signaled, to me anyway,
that this was not a “real dacnis”. The Burns Lab genetic data show that it is distant
from true Dacnis, and clusters with other high-elevation neo-tanagers
like Diglossa and Catamenia.
BLI, as per their usual policy, kept the parental name with the nominate
form despite the fact that it has the much smaller distribution. By our SACC guidelines on
English names,
either petersi would retain the parental name because of its wider
distribution, or new names should be created for both. Then, there is the potential issue of whether
to retain “Dacnis” at all. The English
name Dacnis would have a helpful 1-to-1 match with the genus name, which is
currently hard to do in Thraupidae, if we changed Xenodacnis to
something like “Xenodacnis” as the “last” name, which would retain the former
connection but emphasize the “foreign” angle.
BLI’s “Streaked Dacnis” implies to most that it is just another one of
the real Dacnis and is unacceptable in my opinion; even an awkward “Streaked
Tit-like Dacnis” with a comparable modifier for parina s.s. would be
preferable in my opinion. Dan Lane
suggested to me the potential group name “Tit-dacnis”. Anyway, let all that incubate while we
deliberate on species limits. I’m all
for stability, but if we destabilize species limits, then that would also be
the propitious time to consider a new group name. By the way, Dacnis itself is nothing more
than an alphabetical barcode in terms of a meaningful name – from Jobling: “daknis type of bird from Egypt, otherwise
unidentified, mentioned by Hesychius and Pompeius
Festus.
References: (see SACC
Bibliography
for standard references)
AGUILAR, J. M. 2019. Análisis de la distribución geográfica del género Xenodacnis
(Aves: Thraupidae) utilizando el
modelado de nicho ecológico. Revista
Peruana de Biología 26: 317-324.
AGUILAR, J. M. 2021.
Morphological variation in the Tit-like Dacnis (Xenodacnis parina):
a call to revise the taxonomic status of Ecuador’s population. Ornitologia Neotropical 32: 51-55.
BOESMAN, P. 2016m.
Notes on the vocalizations of the Tit-like Dacnis (Xenodacnis parina). HBW Alive
Ornithological Note 393. In: Handbook of the Birds of the World Alive. Lynx
Edicions, Barcelona. https://doi.org/10.2173/bow-on.100393
BOND, J. 1955.
Notes on Peruvian Coerebidae and Thraupidae. Proceedings Natural Academy Sciences
Philadelphia 107: 35–55.
GEORGE, W. G. 1964. Rarely seen songbirds of Peru’s high
Andes. Natural History 78: 25-29.
Van Remsen, July 2024
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Niels Krabbe (voting for Remsen): “NO. I mustered enough energy to compare all
the 105 different Xenodacnis recordings in XC and ML (25 "cajaensis" from Azuay, Ecuador, 4 bella
from Amazonas and Cajamarca, 54 petersi from W Cajamarca, Ancash, and
Lima, 9 "arequipae" from Arequipa, and 13 nominate parina
from Junín and Cuzco), 38 of which were added to libraries since Boesman's 2016
analysis.
“I
also sorted the measurements given by Aguilar (2021) and compared them with
body masses of birds I collected in Ancash and Apurímac, and concluded that
size is extremely variable (as I had also noted for Cranioleuca baroni
in Ancash and Lima) and not a very useful character, when defining taxa of Xenodacnis.
This leaves bella characterized only by the lesser extent of blue on the
crown in two females when compared with three females of petersi (Bond
1955), in my opinion rather too small a sample size to accept bella as a
valid taxon. That bella should be intermediate in size between parina
and petersi is not corroborated by the measurements at hand, rather on
the contrary. Similarly, the supposed characters defining "cajaensis" do not seem to stand up to scrutiny.
The pale brown vent of the female ("ferruginous vent in all X. parina
subspecies from Peru") looks much like vents of females of bella
and petersi posted in this proposal. As for size difference of "cajaensis", a male I collected in Ancash
(MHNJP, tissue ZMUC 116093) was as heavy (23 g) as in the heaviest male of
"cajaensis". So as to morphology and
plumage, only two taxa seem to be valid: the rather small nominate parina
and the streaked petersi.
As
to the vocal analysis, Boesman's reason for placing the Arequipa birds with
nominate parina was based on vocalizations (not distribution as stated
in the proposal). I concur with Boesman, that the majority of recordings of
"arequipae" are most similar to those of nominate parina,
the only exception being one recording by Ted Parker (ML33844), which is of the
raspy notes so typical of petersi, "bella" and "cajaensis". So, although "arequipae"
has plumage like petersi, "bella" and "cajaensis", it is vocally intermediate with
nominate parina.
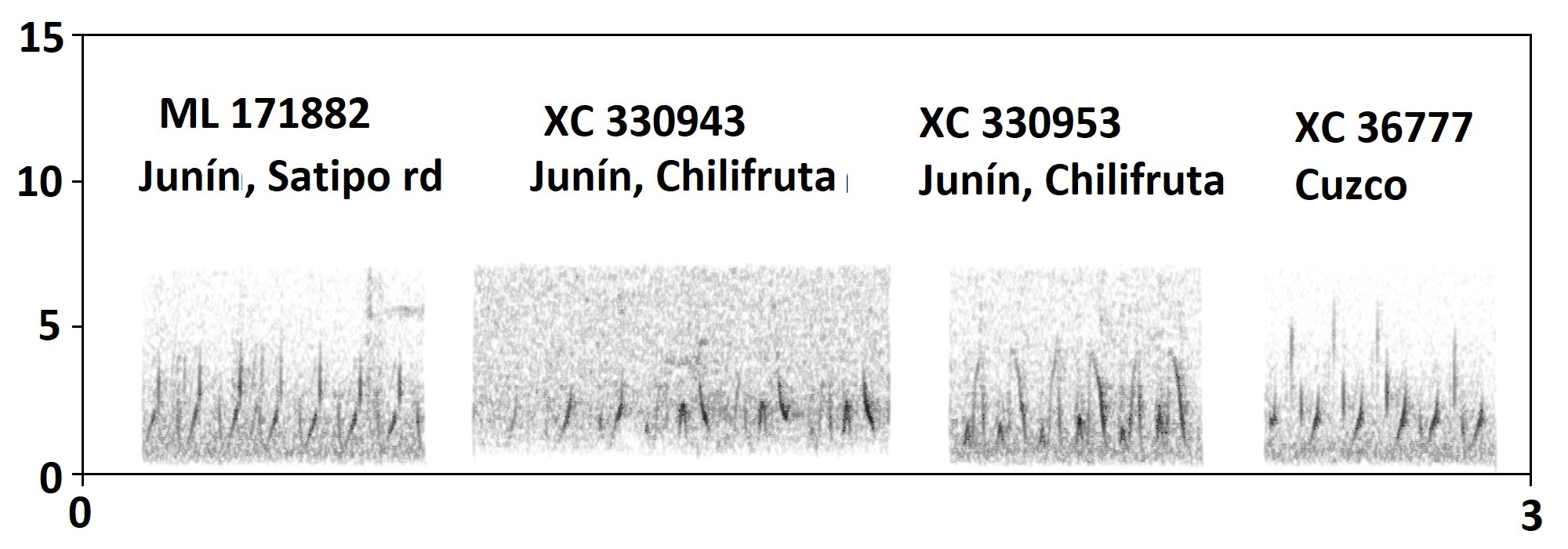
“Another
critical recording is XC330954, uploaded 12 August 2016 and evidently not
analyzed by Boesman. It was recorded by Dan Lane above Chilifruta,
Junín, i.e. nominate parina by range. It includes clear whistled song
typical of petersi, "bella" and "cajaensis".
“These
two cases of intermediacy lead me to believe it highly likely that any two
forms would interbreed, if they were to come into contact. Whether they are
likely genetically incompatible (i.e. diverged more than 2.5 my ago), remains
to be shown by future analyses.
“On
present knowledge, I must vote NO to the proposal.”
Comments from Areta: “NO, but narrowly so, until someone
amasses more carefully vetted data (and hopefully gets more recordings and
genetic data). This seems like a mess with several holes to me. The Ecuador, petersi-bella,
Arequipa, and parina
populations seem to have different songs. Overall, the main plumage and song
types can be grouped in two clusters Ecuador, petersi-bella, Arequipa on
one side and parina on the other.
However, the complex vocalizations have not been well characterized by Boesman
(2016) or Aguilar (2016). Note for example that some recordings from Ancash (petersi) show long and fast
songs, while some recordings from Arequipa also show vocalizations that do not
fit nicely with the characterization by Boesman (2016) (see for example the
spectrogram F in Aguilar 2016]). Aguilar (2016) also indicated that the
Ecuadorian populations deserve subspecific recognition (he coined the name cajaensis,
but to my knowledge this has not been adequately published) and reports on
their plumage and vocal distinctions. The taxon bella has been considered as somewhat
intermediate between parina
and petersi (in
size and male plumage) by Zimmer (1942) despite its geographic position and it
remains vocally little recorded, although how little is known indicates
affinity to petersi
(Van´s photographs of females seem to provide some support on this
intermediacy). The Arequipa population seems also widely allopatric to the petersi populations to which
it is supposedly allied, and also seems to differ from them vocally: do they
also differ in plumage? Aguilar (2016) erroneously included this population in parina as well.
“I think that there is room for a rigorous study here
(those by Aguilar 2016, 2019, 2021 are a beginning). If we were to start from
scratch, I would certainly endorse a 2-species split based on plumage, and this
was also Zimmer´s (1942) preference.
“In our WGAC discussions Tom Schulenberg clarified
that the distributions are not as easy as often assumed. In his own words:
"I just want to point out
that the 2016 vocal analysis by Boesman mischaracterizes the distributions of
the two groups. (this isn't Boesman's fault, he's simply following the original
Hilty account in HBW, which also got this wrong). Boesman assumes that the
pattern is one of simple north/south replacement: northern petersi/bella
and southern nominate parina, but this being Peru, that's not quite the
way it works. Petersi and bella generally occur farther north, on
the west slope of the Andes and on both sides of the interior Marañón Valley.
but the population on the west slope of the Andes in southern Peru in Arequipa
also is of the northern type (presumably petersi), even though it
appears to be highly disjunct from the closest (known) population of petersi
in northern Lima. the Arequipa population is very poorly known, and it may be
that the only specimens in the US are one or two that William George collected
in 1962. I've seen this material (American Museum of Natural History) and
should have photos somewhere, which I'll try to locate. there may well be
additional specimens in the collection in Arequipa, and perhaps we can request
photos of these if need be. Anyway, the upshot is that nominate parina
of course occurs in southern Peru, but only in the interior valleys, and very
locally on the east slope as well; and while petersi and bella
primarily occur in northern Peru, there is an outpost on the west slope in
southern Peru in Arequipa.’
“Also,
regarding the Arequipa populations, Tom provided the following evidence. Three females. from left to right, these are
from Ayacucho (parina),
Lima (presumed petersi),
and Arequipa (Photo courtesy of T.S. Schulenberg):

“Five
males. from top to bottom, these are from Ayacucho (parina), Cusco (parina), Arequipa, Arequipa, and Lima (presumed
petersi) (Photo
courtesy of T.S. Schulenberg)

“Also
the Aguilar (2016) reference:
Aguilar, JM (2016) Description and Conservation
status of a new subspecies of Xenodacnis parina (Aves:
Thraupidae) from the Ecuadorian Andes. M.Sc. thesis, Pontificia Universidad
Católica del Ecuador, Quito, Pichincha, Ecuador.
Additional
comments from Niels Krabbe: “After a closer examination of
Arequipa recordings, I do find them all closer to petersi than I did at
first. This leaves only the Chilifruta recordings
intermediate.”
Comments
from Stiles:
“NO, for now. The series of photographs of plumages presented here rather
strongly suggest that two species could be recognized: petersi of
Ecuador-N Perú-Arequipa, and parina of S Perú. However, the vocal data
may muddy the picture (but note that Arequipa dept. actually extends up to what
looks like the E-W divide of the Andes here and might not match with the
“interior valleys” cited for parina. Hence, while I lean toward
recognizing two species, a more thorough analysis of the vocal data showing the
spectrograms would clinch (or not) the case.”
Comments
from Lane:
“NO. While I suspect that a split or two will be in order once we have a better
handle of the patterns among these taxa, I think we really need much better
evidence to act upon. At present, what we have simply isn’t clear enough.”
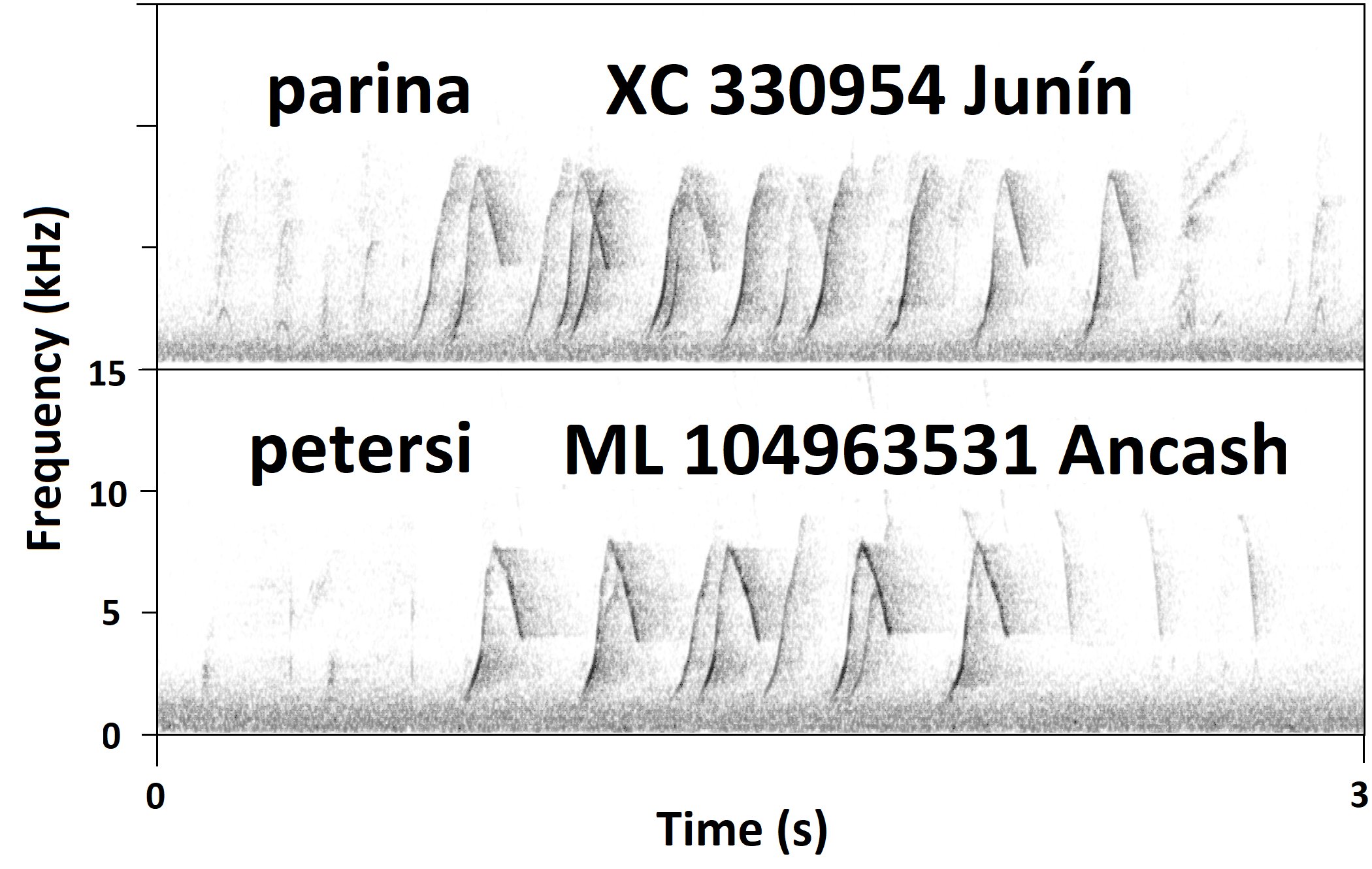
Additional
comments from Niels Krabbe: “Here a spectrogram of the
critical Chilifruta recording of parina
compared with an Ancash recording of petersi, to show the similarity:”
“Here a spectrogram of the bubbly series
only known from parina.”:
Comments
from Robbins:
“NO. I agree with others, especially the analyses
done by Niels, that there needs to be clarification on both morphology and
vocalizations before we do any splits. Genetic
data may also help elucidate on whether more than one species should be recognized.
So, for now, I vote NO treating petersi
as a species.”
Comments
from Claramunt:
“YES. The rationale for voting Yes in the proposal is spot on. There are two
forms here with substantial plumage differences, and they were lumped without
providing ANY evidence. The fact that there are few specimens in museums and
there is additional variation across populations made past taxonomists a bit
doubtful. But the plumage differences stand clearly when examining new specimen
material and photographic records too. The Arequipa population is clearly petersi.
I think that we should revert the unjustified lumping and the burden of proof
should be on those trying to demonstrate that these two taxa are conspecific.”
Comments
from Zimmer:
“A very tentative YES, based mainly on the premise that there are two
phenotypically distinctive populations that were originally treated as
specifically distinct, and then lumped without justification. Even if there are some holes in the data
sets, and a few intermediate specimens, I agree with Santiago that the onus
should be on those advocating for the lump.
But, as I said, I don’t feel strongly about it one way or the other – it
has been some time since I’ve encountered these birds in the field, so it’s not
a group that I pretend to know well in life.”
Comments
from Bonaccorso:
“NO for now. I love the argument about the lack of a justification for lumping
all the forms in one species; it is a tempting place to start the discussion.
However, when the specimens are lined up as in the “Five males” figure provided
by Nacho, where there is a clear cline in ventral plumage, it is difficult to
defend that parina is very different from petersi and bella.
Also, judging from the spectrograms shown by Niels, the songs of parina
and petersi are very similar. Finally, we have no genetic data to
support the split.”
Comments
from Anna Hiller (voting for Del-Rio): “NO - (for now) the presence of intermediate
recordings and potentially unexamined specimens from populations with more intermediate
phenotypes leads me to conclude that a more thorough assessment is sorely
needed (with additional recordings and genetic data). The Aguilar work did a
good first-pass of examining morphological / size data and ecological niche,
making me even more intrigued to see additional data, especially quantitative
color data (female crown color, and an assessment of variation in steaking in
the males). I agree that the Ecuador individuals represent a distinct
population, but whether they rise to the level of species or not remains to be
seen.”