Proposal (1024) to South
American Classification Committee
Treat Rhegmatorhina
berlepschi (Harlequin Antbird) as conspecific with R. hoffmannsi
(White-breasted Antbird)
Background:
The
genus Rhegmatorhina currently comprises five mostly allopatric species,
historically treated as separate species taxa based on diagnostic differences
in plumage (Cory and Hellmayr 1924, Zimmer and Isler
2003). Most are similar in behavior, and some taxa considered separate
species, such as R. berlepschi and R. hoffmannsi, are
undifferentiated vocally (Willis 1969). Although
differences in vocalizations are considered the major phenotypic marker for
species differences in antbirds, plumage differences may be important in
antbirds with ornamented plumages or conspicuous, colorful bare skin patches,
such as Rhegmatorhina (Skutch 1996, Zimmer and
Isler 2003). Genetic work documented structured mitochondrial genotypes
congruent with taxonomic boundaries, but shallowly diverged nuclear genotypes (Ribas et al. 2018, Harvey et al. 2020). The classification of the genus into five species seemed
reasonable given the information we had until now.
New
information:
Recent
genetic research revealed extensive nuclear introgression between R. berlepschi
and R. hoffmannsi across hundreds of kilometers (Del-Rio
et al. 2022). The data is mostly consistent with neutral diffusion of
their nuclear genomes, although narrower
mitochondrial clines imply some selection maintaining mitochondrial
differentiation (Del-Rio et al. 2022). That
berlepschi and hoffmannsi hybridize extensively despite differences
in plumage (Del-Rio et al. 2022) indicates
that their plumage differences have little (if any) impact on reproductive
isolation. In summary, this data indicates weak overall reproductive isolation
between berlepschi and hoffmannsi.
Recommendation:
Given
the evidence of free nuclear gene flow between berlepschi and hoffmannsi
(Del-Rio et al. 2022), their taxonomic status
warrants reconsideration. The hybrid zone between these taxa is significantly
wider than those between many bird lineages considered conspecific (Price 2008). The correct taxonomic treatment seems
straightforward in light of the new evidence: berlepschi and hoffmannsi
are weakly reproductively isolated and, therefore, should be treated as
conspecific. The name hoffmannsi (Hellmayr, 1907) has priority over berlepschi
Snethlage, 1907, by some months.
References
Cory, C.
B., and C. E. Hellmayr (1924). Catalogue of birds of the Americas and the
adjacent islands in Field Museum of Natural History. Volume XIII, Part III.
Field Museum Press.
Del-Rio,
G., M. A. Rego, B. M. Whitney, F. Schunck, L. F. Silveira, B. C. Faircloth, and
R. T. Brumfield (2022). Displaced clines in an avian hybrid zone
(Thamnophilidae: Rhegmatorhina) within an Amazonian interfluve.
Evolution 76:455–475.
Harvey, M.
G., G. A. Bravo, S. Claramunt, A. M. Cuervo, G. E. Derryberry, J. Battilana, G.
F. Seeholzer, J. S. McKay, B. C. O’Meara, B. C. Faircloth, S. V Edwards, et al.
(2020). The evolution of a tropical biodiversity hotspot. Science
370:1343–1348.
Price, T.
(2008). Speciation in birds. Roberts & Company.
Ribas, C.
C., A. Aleixo, C. Gubili, F. M. d’Horta, R. T. Brumfield, and J. Cracraft
(2018). Biogeography and diversification of Rhegmatorhina (Aves:
Thamnophilidae): Implications for the evolution of Amazonian landscapes during
the Quaternary. Journal of Biogeography 45:917–928.
Skutch, A.
F. (1996). Antbirds and ovenbirds: their lives and homes. University of Texas
Press.
Willis, E.
O. (1969). On the behavior of five species of Rhegmatorhina,
ant-following antbirds of the Amazon basin. The Wilson Bulletin 81:363–395.
Zimmer, K.
J., and M. L. Isler (2003). Family Thamnophilidae (Typical Antbirds). In
Handbook of the birds of the world (J. del Hoyo, A. Elliot and D. A. Christie,
Editors). Lynx Edicions, pp. 448–681.
Rafael
Lima, July 2024
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Stiles:
“YES. This is an easy choice, with a solid publication based upon abundant and
appropriate data - in contrast to all those recent ones to evaluate splits
based on fragmentary, incomplete and often conflicting pieces of data!”
Comments
from Lane:
“NO. Glaucia’s study is an intriguing one, but even she didn’t propose lumping
the two taxa because of the hybrid zone. The hybrid zone seems to be moving,
and doesn’t cover a large area (compared to the Myioborus case in Prop
1003, for example), suggesting that it is not a static system, but rather a
dynamic one that may end with one form swamping out the other.”
Comments
from Robbins:
“NO. As we have seen over and over, mitochondrial
data has limited utility and can mislead interpretations of taxonomy. Given the
nuclear data coupled with the reported lack of differentiation in primary
vocalizations, this seems to be rather straightforward in that these two should
be considered conspecific.”
Comments from Areta: “NO. I was waiting for
Glaucia´s input on this. The genetic situation is not as simple as described in
the proposal, and there are several nuances discussed in the meaningful paper
by Glaucia and collaborators. Meanwhile, in the face of a proper study measuring
vocalizations of Rhegmatorhina,
I decided to take a listen. The vocal similarities between hoffmannsi and berlepschi are striking and obvious even without
a quantitative study (and in agreement with the broad hybridization area found),
but what caught my attention are the also very similar vocalizations of gymnops (east of the Tapajós): songs and the
burry/churring calls sound pretty much identical. Glaucia´s work has also shown
a shallow divergence between gymnops and hoffmannsi/berlepschi.
Even if we know of no intergrades between berlepschi and gymnops (or is there any evidence for this?),
the seeming lack of vocal differences in their primary songs, minor plumage
differences (akin to those between berlepschi and hoffmannsi),
and shallow genetic divergence (see haplotype network in Del-Rio et al. 2022)
all would seem to indicate that, if east-Tapajós gymnops is given the chance to interact
reproductively with west-Tapajós berlepschi,
they would hybridize freely. Then I remembered a possibly relevant work and
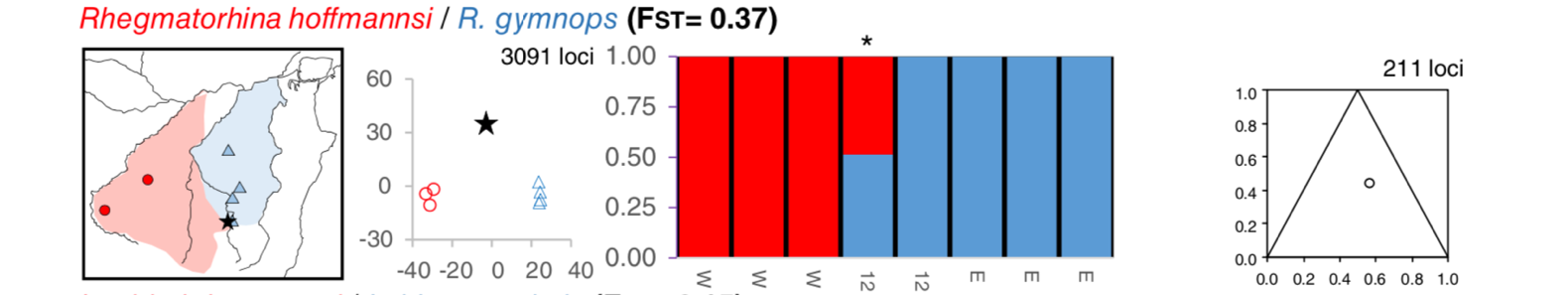
found it: Weir et al. (2015; Evolution 69-7: 1823–1834) have found
hybridization close to the Tapajós headwaters (the black star in the figure
shows a hybrid between "hoffmannsi" and gymnops, I use quotation marks as it
is not clear to me how these birds looked like and what would happen with
phenotypes in that area):
“Weir et al. (2015) also add that "Mitochondrial sequence based estimates
of gene coalescence times between parental populations from the Rondonia and
Tapajos endemism centers ranged from just 0.7 Ma in L. iris / L.
nattereri and R. hoffmannsi / R. gymnops to 4.1 Ma in W.
poecilinota / W. vidua (Fig. 4)."
and "Ironically, the
taxon pairs with the most clearly differentiated plumage patterns, and which
have always been recognized as specifically distinct (R. hoffmannsi/R.
gymnops; L. nattereri/L. iris), are also the
youngest (<1 Ma) in our sample."
The discussion of Willis (1969) provides interesting
perspectives and somehow prepared the scenario for the modern findings: "The forms gymnops, berlepschi, and hoffmannsi are practically identical in behavior.
Oddly, berlepschi looks like a hybrid between gymnops to the east and hoffmannsi to the west."
And more:

“Finally, note that gymnops
was described by Ridgway in 1888, so that name would have priority
over berlepschi
and hoffmannsi. It seems
to me that a 3-way lump would be a reasonable treatment, but I would like to
hear from those with extensive Amazonian experience (I am eagerly waiting for
Bret´s and Kevin´s surely encyclopedic experience on this group). Even if at
present the Tapajós is impassable, I would like to see a deeper genetic and
vocal study in order to change our current taxonomic treatment. It seems that
the current 3-species treatment of the east-Madeiran
Rhegmatorhina is wrong,
but how many species should be recognised is debatable.”
Comments from Robb Brumfield (who has Jaramillo vote): “NO.
Hybridization between hoffmannsi
and berlepschi
is clearly extensive. The center of the hybrid zone is composed
almost entirely of recombinant individuals, and introgression of alleles
extends deep into the distributions of the two hybridizing taxa. These patterns
are typical of most avian hybrid zones for which we have genome-scale data.
However, Del-Rio et al (2021) found five diagnostic SNPs that fully
distinguish the two taxa and, using those, identified two individuals from the
hybrid zone as putative F1s. Both individuals were heterozygous at all
five diagnostic nuclear SNPs, and both had intermediate plumage. The presence
of these F1s is atypical
of most avian hybrid zones. The two F1s suggest that
strong reproductive
barriers exist between the two taxa in some parts of their nuclear genomes.
There are of course nuances to hybrid zone models that can change
interpretations based on assumptions, but it may be that strong selection on a
few nuclear loci is sufficient to maintain nuclear divergence between
hoffmannsi and
berlepschi. Whether the presence of strong reproductive
barriers at a few nuclear loci is sufficient evidence for species status is
unclear to me. Weir et al. (2015,
Evolution 69:1823)
found evidence of hybridization between
R. hoffmannsi and
R. gymnops where they
come into contact. My take is that
Rhegmatorhina is
reminiscent of Manacus
manakins, in that narrow hybrid zones occur wherever species come
into contact. Based on the data in
Del-Rio et al. (2015) and the fact that
Rhegmatorhina hybridization
and speciation is the focus of ongoing research, my vote is to not make any
taxonomic changes now.”
Additional comments from Rafael Lima: “I agree with Areta that a three-way lump
seems like the most reasonable approach, making gymnops the oldest name. That option initially escaped my
attention, and I thank Juan for pointing it out.
“I really
appreciate Robb’s thoughtful input on this matter and would like to offer some
additional reflections based on his comments.
“Robb mentions
that Rhegmatorhina hybrid zones are reminiscent of those in Manacus, where narrow hybrid zones occur wherever species come
into contact. However, I am not entirely convinced that the hybrid zone between
hoffmannsi and berlepschi can
be accurately described as "narrow." Of course, the terms
"narrow" and "broad" are subjective, but if the Rhegmatorhina hybrid zone is considered narrow, I
wonder what a "broad" hybrid zone would look like. The hybrid zone
between
hoffmannsi and berlepschi is, in fact, wider than many hybrid zones
typically referred to as "wide" in the literature, including those
between bird lineages considered conspecific (e.g., Price 2008).
“I completely
agree with Robb's point regarding whether strong reproductive barriers at a few
loci should suffice for species status when there is otherwise free gene flow
throughout the genome. I don’t have an answer for that question. However,
regardless of what the preferred cutoff should be, I believe this case may set
a precedent for future proposals. In the interest of consistency, those voting
"no" on this proposal might need to consider whether they would also
support splitting the many bird taxa currently classified as subspecies that
have similar hybrid zones and reproductive isolation levels. A "no"
vote might imply that bird lineages forming hybrid zones several hundred
kilometers wide, dominated by late-generation hybrids, should be considered
separate species under the BSC because of reproductive barriers at a few loci
amid widespread introgression elsewhere across the genome.
“I wonder why two
vocally undifferentiated antbird lineages that hybridize extensively over
hundreds of kilometers (i.e., berlepschi and hoffmannsi) should be regarded as separate
biological species while other similar antbird lineage pairs should be
considered conspecific. For instance, I recall the Thamnophilus capistratus proposal (Proposal 890). Why should Thamnophilus doliatus capistratus and the rest of Thamnophilus doliatus be considered conspecific when they may
have a hybrid zone less than 100 km wide, while Rhegmatorhina taxa with substantially wider hybrid
zones should be considerate separate species? The Thamnophilus capistratus proposal is merely an example that came
to mind, but there are numerous similar cases within the Thamnophilidae. While
I no longer think that capistratus should
necessarily be treated as a separate species, I do believe that consistent
application of criteria is crucial. If criteria are to be applied consistently
across taxa, we may need to reconsider species limits in many antbird taxa
based on the precedent potentially set by Rhegmatorhina.
“As a final note,
I personally view species limits as hypotheses formulated within the context of
currently available evidence. This is how I understand science to work, or at
least how it should ideally function, in my opinion. We should make decisions
based on the evidence at hand, not on potentially different evidence that may
emerge in the future. Therefore, I am not entirely convinced by the argument
that taxonomic changes should be postponed due to ongoing research on Rhegmatorhina hybridization. If we avoid taxonomic
changes because of ongoing research, we might never make any changes. I believe
the decision here should be as straightforward as the recent NACC decision to
treat Corvus caurinus as conspecific with C. brachyrhynchos, for example. Common to both cases is the existence of recent
studies providing crystal clear evidence for extremely weak reproductive
isolation between the two lineages involved. If Del-Rio et al. (2021) did not
provide sufficient evidence of weak reproductive isolation between berlepschi and hoffmannsi, I wonder what future research could
possibly demonstrate conspecificity between these two taxa. Should that
evidence arise, we can adjust accordingly, as we scientists routinely do.”
Comments from Claramunt: “Tentative NO. I was going to
vote Yes, persuaded by Rafael’s proposal and a cursory examination of Glaucia’s
superb* paper. R. berlepschi and hoffmannsi are clearly
hybridizing and forming a relatively wide hybrid zone with intergradation of
traits, and admixture of genomes. Songs are the same and the plumage
distinction didn’t impede interbreeding. It seems that there are no barriers to
interbreeding. I’m not convinced by the argument about the purported
identification of F1 individuals. In a sample of 222 individuals and only five
diagnostic loci, it would not be hard to find a handful of specimens that look
like F1s by chance, just by being heterozygotes for the five loci. Unless I’m
missing something, I don’t see how this method would lead to correct
identification of F1s. And it seems that it would be impossible for true F1s to
be formed anyway given that there are no parental pure forms in geographic
proximity.
“The unavoidable conclusion under the biological species concept
is that these two taxa should be lumped. Many recent proposals for species
status are being rejected because of the speculation of potential interbreeding
in hypothetical sympatry. Well, here there is actual evidence of widespread
interbreeding and no signs of reproductive barriers.
“Having said that, it is possible that the degree of genomic
admixture is being overestimated. Each SNP has very little information about
genealogy, and at least some methods used to analyze SNPs do not take a
historical/evolutionary perspective, e.g. allele A suggests ancestry X if A is
frequent in X, regardless of whether A is the ancestral state or not. If allele
A is ancestral, sharing it does not indicate close relationships in the
genealogical network (basic phylogenetics applied to genealogies). I’m going to
do some research to see if I can back up my suspicion with some evidence. In
the meantime, I feel entitled to suspect that the width of the SNP clines and
the degree of admixture is being overestimated. This may be a case of a, maybe
not narrow but relatively constrained hybrid zone (200km?) on genomic
backgrounds that are differentiated. Then the question would be whether the
hybrid zone is a tension zone maintained by some sort of selection against
hybrids or assortative mating. Is the hybrid zone stable or moving? Is it
shrinking or expanding? There may be room for still considering these two taxa
separate species even if hybridizing. For that reason I tentatively vote NO,
waiting for more comments and discussion about this case, that may set a
precedent for decisions based on this kind of scenario.”
Comments from Del-Rio: “NO. It is a complicated case,
primarily because most of the cases we analyze here at SAAC are based on
plumage, song, morphology, a few genes, or—in the best-case scenarios—reduced
representation genomic data (usually less than 1% of whole genomes). In the
case of Rhegmatorhina, however, we have all of the above plus a
high-quality whole genome and whole genome resequencing data at a decent
coverage for almost one hundred individuals.
“In the era of whole genomes, let’s see what happens. How are we
going to define what constitutes strong versus weak levels of reproductive
isolation?
“The results and ideas in the Evolution paper (Del-Rio
et al., 2022) have evolved over the last couple of years. I won’t go into
details here, but my most recent results show: (1) several genomic islands of
differentiation between Rhegmatorhina hoffmannsi and R. berlepschi;
(2) strong differential selection indicating assortative mating on regions of
the genome involved in the definition of plumage coloration (Del-Rio et al in
prep.).
“I vote NO because: (1) The argument that the Rhegmatorhina
hybrid zone is too wide is flawed, as the hybrid zone width varies depending on
which part of the genome is being analyzed. Some regions of the genome and some
plumage features show narrow and steep clines. That’s the beauty of the
system—a unique opportunity to understand which parts of the genome are
preventing species fusion; (2) Additionally, with the available methods, the
complex geography of the system leads to an overestimation of cline widths; (3)
I am still working to understand how much these islands of genomic
differentiation translate into reduced fitness in hybrids; (4) I am still
trying to perform experiments to test for assortative mating based on plumage
and vocal characters; (5) I am still analyzing vocal and environmental
variation between the two forms; (6) In Brazil, “subspecies” status is rarely
considered in conservation efforts, and R. berlepschi might become a
species of concern with the advance of deforestation in Amazonia.
“Regarding the idea of gene flow with R. gymnops, my data
does not corroborate the findings of Weir et al. (2015) so far. Again, this is
another aspect I am still investigating.”
Comments from Del-Rio: “NO. It is a complicated case,
primarily because most of the cases we analyze here at SAAC are based on
plumage, song, morphology, a few genes, or—in the best-case scenarios—reduced
representation genomic data (usually less than 1% of whole genomes). In the
case of Rhegmatorhina, however, we have all of the above plus a
high-quality whole genome and whole genome resequencing data at a decent
coverage for almost one hundred individuals.
“In the era of whole genomes, let’s see what happens. How are we
going to define what constitutes strong versus weak levels of reproductive
isolation?
“The results and ideas in the Evolution paper (Del-Rio et
al., 2022) have evolved over the last couple of years. I won’t go into details
here, but my most recent results show: (1) several genomic islands of
differentiation between Rhegmatorhina hoffmannsi and R. berlepschi;
(2) strong differential selection indicating assortative mating on regions of
the genome involved in the definition of plumage coloration (Del-Rio et al in
prep.).
I vote NO because: (1) The argument that the Rhegmatorhina hybrid
zone is too wide is flawed, as the hybrid zone width varies depending on which
part of the genome is being analyzed. Some regions of the genome and some
plumage features show narrow and steep clines. That’s the beauty of the
system—a unique opportunity to understand which parts of the genome are
preventing species fusion; (2) Additionally, with the available methods, the
complex geography of the system leads to an overestimation of cline widths; (3)
I am still working to understand how much these islands of genomic
differentiation translate into reduced fitness in hybrids; (4) I am still
trying to perform experiments to test for assortative mating based on plumage
and vocal characters; (5) I am still analyzing vocal and environmental
variation between the two forms; (6) In Brazil, “subspecies” status is rarely
considered in conservation efforts, and R. berlepschi might
become a species of concern with the advance of deforestation in Amazonia.
“Regarding the idea of gene flow with R. gymnops, my data
does not corroborate the findings of Weir et al. (2015) so far. Again, this is
another aspect I am still investigating.”
Additional comments from Rafael Lima: “Considering the comments above, it seems
unlikely that any amount of evidence or argument will alter the trajectory of
this proposal. Nevertheless, I would like to share some final thoughts:
“1. My impression
from all this is that there is a strong reluctance to change, as most arguments
for maintaining the current classification hinge on predictions about ongoing
studies or conservation-oriented reasoning, rather than drawing much more straightforward
conclusions from the available evidence. While I’ve already expressed my
thoughts on this, I feel it’s important to reiterate that taxonomic decisions
should be grounded in the evidence we currently have, not on what future
research might reveal. I fully understand the desire for taxonomic stability,
especially for those working in downstream fields that use checklists like
this. However, prioritizing stability over scientific rigor severely undermines
taxonomy as a discipline. I therefore urge that decisions be made based on
present evidence rather than futurology.
2. The conclusion
that these taxa should be considered conspecific appears so evident in the
results of Del-Rio et al. (2022) that I initially thought this proposal would
be straightforward. However, based on the comments above, it seems I may not
have fully elaborated on my interpretation of these results. One key point I
thought would be clear to anyone reviewing the results from Del-Rio et al.
(2022) is the modality of the hybrid zone. The modality of a hybrid zone is a
critical indicator of the strength of reproductive isolation between two
interbreeding populations (nicely reviewed in Jiggins & Mallet 2000, Trends
Ecol. Evol. 15: 250-255). In Rhegmatorhina, the hybrid zone is clearly
unimodal. Most importantly, it is late-generation hybrids that dominate. I
can’t think of a more compelling indication that overall reproductive isolation
is very weak in this system. If assortative mating existed, the zone would be
bimodal. If strong postzygotic isolation occurred, through selection against
hybrids or genetic incompatibilities, first-generation hybrids, not
backcrosses, would dominate. Neither assortative mating nor postzygotic
isolation seems to be in play.
“Glaucia mentions
the presence of steep clines, possible assortative mating, and reduced hybrid
fitness as potential reasons to maintain the taxa as separate species. Theory
predicts that as differentiation accrues between any two populations, some
degree of reproductive isolation will almost always be present. Unsurprisingly,
it is not unusual to observe steep clines, assortative mating, and reduced
hybrid fitness within species. The critical question when applying the
BSC is not whether these phenomena are at play, but whether they are strong
enough to prevent the fusion of the populations involved. In the case of Rhegmatorhina,
there are some narrow clines, but they exist alongside many broader ones. Are
these narrow clines causing moderate or strong overall reproductive isolation?
The evidence suggests not. Regarding assortative mating and hybrid fitness, the
unimodal nature of the hybrid zone makes it implausible that either is
occurring at a significant level. It is not entirely clear to me how there could
be assortative mating given that there are no parentals in the hybrid zone, or
how hybrids could suffer from reduced fitness when backcrosses are so
predominant. Even if we assume some degree of non-random mating and reduced
hybrid fitness, the current evidence strongly suggests that these putative
barriers are insufficient to prevent extensive gene flow. Gene flow remains
abundant, despite any potential barriers. The way I see it, the burden of proof
is clearly on the current multiple-species treatment, for which the evidence
has long been weak (as noted by Willis 1969).
“Finally, it is
not true that subspecies taxa are not considered in conservation efforts in
Brazil. The Brazilian Red List of Threatened Species does include
subspecies-level assessments for birds. However, this point should not even be
a factor in this discussion, as the decision here should be independent of
conservation concerns. (Taxonomic decisions motivated by conservation have
already been addressed in other proposals, so I will refrain from elaborating
further.) Otherwise, species limits in Rhegmatorhina will, after this
voting, be based more on speculation and conservation-oriented reasoning than
on existing evidence.”
Additional comments from Claramunt: “I change
my vote to YES. At the end of the day, I’m with Rafael on this. The evidence of
introgressive interbreeding is just overwhelming. There are no signs of
selection against hybrids: the paper concludes that neutral diffusion cannot be
rejected. There are some hints of “islands of differentiation” and reduced
hybrid fitness in some part of the genomes, but those are clearly not
sufficient as reproductive barriers and genes are profusely flowing in both
directions. The conclusion is inescapable under any species concept:
reproductive isolation is nonexistent, diagnosability is broken, lineages are
intertwined.
“It doesn’t get better than that when it comes to evidence of
interbreeding. It is not only the absolute width of the hybrid zone what
matters but how broad it is in comparison with the rest of the distribution of
the species involved. In this case, the tail of the hybrid zone gets so deep
into the berlepschi territory that there are barely any pure berlepschi
anymore (see the ancestry analysis in Fig. 3). The paper by Glaucia et al. is
superb, and we are in a period in which we need to pay close attention to
analyses of genomic data. The genomic revolution would likely change our
perspectives on how we delimit species and even on what species are. But today,
under current species concepts, the evidence in this case is conclusive.
Keeping berlepschi and hoffmannsi as separate species in the face
of seemingly free introgressive hybridization would be a radical departure from
a century of application of the biological species concept. If our mission is
to produce a coherent and evidence-based classification, we can’t maintain berlepschi
and hoffmannsi as separate species. There is no better evidence of
interbreeding than genomic analyses like this. Plumage and song differences and
similarities, and playback experiments, can suggest reproductive compatibility,
but never prove it. We can’t ignore the best evidence in this case and at the
same time lump other taxa based on much lower quality evidence, in many cases,
educated speculation about what birds would do if in sympatry. Is this a
breaking point in our approach to species delimitation? Actual evidence of
introgressive hybridization does not count anymore? We must surrender to the
evidence if we are to produce a scientific classification.”
Comments from Remsen: “NO. I have looked at the series
assembled by Glaucia and colleagues from the contact zone. Although there are some obvious hybrids,
there are also at least as many phenotypically pure birds from the. contact
zone, and this suggests that mating is not random, thus supporting separate
species treatment under the BSC.”
Comments from Zimmer: “YES, although not an
enthusiastic yes. It pains me, because
these birds are near and dear to my heart, and because both forms are truly
exquisite in their parental phenotypic states.
However, I have examined whole museum trays of obvious hybrids, and the
evidence for a relatively broad hybrid swarm, with no pure parentals of either
type, and apparently, even very few F1 hybrids, appears overwhelming. I’ve gone back and forth on this, given that
Glaucia, who, presumably, knows these birds better than anyone since Ed Willis,
is not advocating for lumping at this time.
On the one hand, lumping hoffmannsi and berlepschi does
solve a conundrum that puzzled Willis and all Rhegmatorhina enthusiasts
since – how, in a group where all of the species have allopatric distributions
separated by major rivers, can two species be found in the same interfluve,
without an obvious geographic barrier, and still maintain isolation? And now, we know, they don’t. Of course, we also now know, from all of the
relatively recent work done in the Aripuanã and Roosevelt drainages, and, to a
lesser extent, along the Teles Pires, that the complex
geologic/hydrographic/hydrologic history of the Madeira-Tapajós interfluve has
resulted in a present-day situation in which relatively small rivers may
present barriers sufficient to maintain isolation and prevent secondary contact
of some understory passerines of low vagility.
That fact, coupled with the complex physical history of the interfluve,
probably provides the answer to Willis’s question: “The relationship of berlepschi and
hoffmannsi is even more puzzling. Both
species occur between the Tapajoz and Madeira, without any large river or
unsuitable habitat to prevent their meeting.
How could two forms or species have evolved?” The one thing that gives me pause over the
decision to lump these two species, is the same thing that I circle back to
when considering similar hybrid zones between several of the larger species of
North American Larus -- how, in
the face of broad hybrid zones, in which mating appears non-assortative and
pure parental types are outnumbered by hybrids, do we explain the maintenance
of pure parental phenotypes over large swaths of the parental ranges? The distributions of hoffmannsi and
(especially) berlepschi are not all that large to begin with. With ongoing regional deforestation eroding
these already small ranges, I would think that in the absence of some
meaningful pre or postzygotic barriers to reproduction, the hybrid zone would
expand to the point of swamping both parental phenotypes. That this has not happened, makes me think
that there are still some factors in play that are promoting isolation, or at
least, slowing allelic penetration from one genome into the other outside of
the hybrid zone. I would like to know if
the hybrid zone is stable or labile, and, if the latter, is the expansion
unidirectional or is it advancing in both directions. Despite all of these reservations, I am,
ultimately, persuaded by the well-reasoned arguments of Rafael and Santiago,
and have to concede that the weight of the evidence at hand leads to the
conclusion to lump hoffmannsi and berlepschi. Conversely, I am not supportive, on current
evidence, of expanding this to include gymnops in a 3-way lump. Despite obvious vocal and behavioral
similarities between the 3 taxa, there is no evidence that I have seen, of a
documented hybrid zone between gymnops and hoffmannsi, even if
Weir et al. (2015) are reporting a single putative hybrid from the narrow
headwater region of the Tapajós. That’s
a far cry in my estimation, from the situation with hoffmannsi and berlepschi. I don’t doubt that gymnops and hoffmannsi
may be only shallowly diverged genetically from one another, but extrapolating
that to support speculation about how the two would behave were they to come
into contact seems premature, given that for the vast majority of their
respective ranges, the two are separated by the length of the mighty
Tapajós. The headwaters region, where
they may contact one another, is also within the notorious Arc of Deforestation
at the southern fringes of Amazonia and would seem an unlikely venue for an
expanding hybrid zone between two terra firme interior-inhabiting
species that are only marginally in contact.
Note Glaucia’s comment that her data, to this point, does not
corroborate the findings of Weir et al (2015) with respect to gene flow with gymnops.”