Proposal (1028) to South
American Classification Committee
Treat Synallaxis
cinerea as conspecific with S. ruficapilla
The
Synallaxis ruficapilla complex currently includes three species taxa:
the Rufous-capped Spinetail (S. ruficapilla), Bahia Spinetail (S.
cinerea), and Pinto’s Spinetail (S. infuscata). Also part of the
complex is a geographically isolated population in the Brazilian state of Mato
Grosso, often cited as a possibly undescribed taxon
(Whitney and Cohn-Haft 2013, Batalha-Filho et al. 2013) but which may merely
represent a range extension of ruficapilla. For a more in-depth
overview of the complex, the reader is referred to the work of Stopiglia et al. (2013).
Phenotype
Stopiglia et al. (2013) examined geographic variation in plumage,
morphometrics, and vocalizations. They found that only two populations in the
complex are phenotypically diagnosable: one corresponding to infuscata and the
other comprising the rest of the complex.
Genotype
Genetic
research identified five mitochondrial lineages within the complex: one
corresponding to infuscata, one to cinerea, one to the Mato
Grosso population, and two lineages within ruficapilla (Batalha-Filho et al. 2013,
2019). All have very shallow nuclear divergence (Batalha-Filho et al. 2013,
2019; Harvey et al. 2020).
Recently, Batalha-Filho
et al. (2019) analyzed two mitochondrial genes, two autosomal nuclear introns,
and three Z-linked genes from many individuals of cinerea and ruficapilla. They discovered a wide hybrid zone between the two near the
Jequitinhonha River. One Z-linked locus showed a narrow cline (~ 28 km), while
all other loci showed wide clines (~ 179–529 km).
In summary, all taxa, including those
that are phenotypically indistinguishable, are diagnosable units with respect
to mtDNA. Additionally, there are other populations with equivalent levels of
mtDNA divergence that are unnamed within the complex, namely the Mato Grosso
population and one population within ruficapilla. The narrow cline for one Z-linked locus suggests some selection on
the hybrid zone between cinerea and ruficapilla, although there is extensive gene flow on the rest of the genome.
How many taxa?
The
S. ruficapilla complex presents a challenging situation for
classification. The
geographic phenotypic variation in the S. ruficapilla complex is similar to that of
several other bird species complexes along the Atlantic rainforest, with a
phenotypically diagnosable populations restricted to north of the São Francisco
River and two somewhat differentiated, but not diagnosable, populations south
of that river (e.g., Lima et al. 2024). In this broader context, S. cinerea seems to be another example of
an avian taxon originally described from a small sample (Pacheco and Gonzaga
1995) that eventually proved undiagnosable with a greater sample (Stopiglia et
al. 2013).
Given that cinerea and ruficapilla are
phenotypically undistinguishable (Stopiglia et al. 2013), the only currently available basis for
recognizing cinerea as a valid taxon (regardless of rank) is
diagnosability on the basis of mtDNA and some Z-linked loci (Batalha-Filho
et al. 2019). Regarding rank,
extensive hybridization between cinerea and ruficapilla suggests that
the current treatment of the two as separate biological species may be in
error.
I recommend a YES vote to recognize as cinerea and ruficapilla as
conspecific, based on the recent evidence of extensive gene flow and the lack
of phenotypic diagnosability between the two.
References:
Batalha-Filho,
H., M. Irestedt, J. Fjeldså, P. G. P. Ericson, L. F. Silveira, and C. Y. Miyaki
(2013). Molecular systematics and evolution of the Synallaxis ruficapilla
complex (Aves: Furnariidae) in the Atlantic Forest. Molecular Phylogenetics and
Evolution 67:86–94.
Batalha-Filho,
H., M. Maldonado-Coelho, and C. Y. Miyaki (2019). Historical climate changes
and hybridization shaped the evolution of Atlantic Forest spinetails (Aves:
Furnariidae). Heredity 123:675–693.
Harvey, M.
G., G. A. Bravo, S. Claramunt, A. M. Cuervo, G. E. Derryberry, J. Battilana, G.
F. Seeholzer, J. S. McKay, B. C. O’Meara, B. C. Faircloth, S. V Edwards, et al.
(2020). The evolution of a tropical biodiversity hotspot. Science
370:1343–1348.
Lima, R.
D., A. C. Fazza, M. Maldonado-Coelho, C. Y. Miyaki, and V. Q. Piacentini
(2024). Taxonomic revision of the Scaled Antbird Drymophila squamata
(Aves: Thamnophilidae) reveals a new and critically endangered taxon from
northeastern Brazil. Zootaxa 5410:573–585.
Pacheco, J.
F., and L. P. Gonzaga (1995). A new species of Synallaxis of the ruficapilla/infuscata
complex from eastern Brazil (Passeriformes: Furnariidae). Ararajuba 3:3–11.
Stopiglia,
R., M. A. Raposo, and D. M. Teixeira (2013). Taxonomy and geographic variation
of the Synallaxis ruficapilla Vieillot, 1819 species-complex (Aves:
Passeriformes: Furnariidae). Journal of Ornithology 154:191–207.
Whitney, B.
M., and M. Cohn-Haft (2013). Fifteen new species of Amazonian birds. In
Handbook of the birds of the world. Special volume: new species and global
index (J. del Hoyo, A. Elliot, J. Sargatal and D. A. Christie, Editors). Lynx
Edicions, pp. 225–239.
Rafael D.
Lima, July 2024
_________________________________________________________________________________________________________
Comments from Claramunt: “YES. I
studied the type series of cinerea at the AMNH myself, and I could not
find any consistent difference with specimens of ruficapilla. The polymorphism
in the song is intriguing, but it is not separating two taxonomic groups, as
demonstrated by Stopiglia et
al. (2012). My impression is
that cinerea should not be recognized even at the
subspecies level; it’s a full synonym of ruficapilla.”
Comments from Robbins: “YES for
reasons outlined in the proposal.”
Comments from Stiles: “YES: the
proposal seems clear and logical given the evidence adduced.”
Comments
from Zimmer: “YES, for reasons nicely summarized in
the Proposal. I do wonder as to whether
Stopiglia et al. (2013) included specimens of the still-undescribed Mato Grosso
birds in their study, and, if so, how many specimens of said population they
examined. Hard for me to imagine that
those birds are not diagnosable, or, that they represent merely a range
extension of ruficapilla – if so, it would not represent just a
significant range extension, but it would also mean that a bird that occupies
Atlantic Forest, including montane forest, over most of its rather wide range,
has a string of isolated populations occupying lowland Amazonian forest-edge
between the Xingu basin and Alta Floresta.”
Comments
from Josh Beck: “Despite not having had time to do an exhaustive
analysis I spent quite some time listening to vocalizations on XC based upon my
memory of the voice of these two taxa being distinctive. I am surprised by the
fact that the vocalizations were not discussed in the proposal and haven’t been
mentioned yet. They are instantly diagnosable by voice. My initial
investigation in Xeno Canto of recordings along the species boundaries didn’t
reveal any intermediate or out of place vocalizations, at least initially
suggesting a clean boundary between the two species based on voice.”
Comments
from Bonaccorso: “YES. Still, I was also surprised that
vocalizations were not discussed in the proposal, especially when many of our
decisions are heavily influenced by song differences. Still, the degree of
hybridization and the lack of diagnostic phenotypic differences seem indicate
that the species-level rank of S. cinerea is not warranted. That said, it looks like a neat system to
study vocal interactions in and out of the contact zone.”
Comment from Rafael Lima: “I
would just like to clarify that vocalizations are addressed in the proposal. It
is explicitly mentioned that Stopiglia et al. (2013) examined geographic
variation in plumage, morphometrics, and vocalizations. As summarized in
the proposal, cinerea was not diagnosable from ruficapilla
in any of the examined phenotypic traits, including vocalizations. I would be
curious to learn more about the specific differences that Josh identified in
the xeno-canto recordings, given that his conclusion that the two taxa are ‘instantly
diagnosable by voice’ contrasts with Stopiglia et al.'s (2013) conclusion.”
Comments from Lane: “NO. I
have to agree with Josh here, and I’m a bit amazed that Stopiglia et al. didn’t
detect vocal differences between S. cinereus and S. ruficapillus…
it actually should be pretty obvious to any listener! Rather than the multi-note
“dddd-dic” with an obvious pause after each song phrase, it’s a more
rapidly-given “d-dic” with very brief pauses or a very rapid series
(d-dic-d-dic-d-dic…) with no pauses at all. If you listen to recordings from
the geographically most proximal populations of S. ruficapillus, they
show no variation overlapping S. cinereus.”
Additional comments from Josh Beck: “Stopiglia et al (2013) basically divided
the vocalization of either species into two phrases – with cinerea
essentially being a one note first phrase and a one note final phrase, and ruficapilla
being a fast series then a final note. They then state “The number of elements
varies geographically, with zero to six elements north of the Jequitinhonha
River and one to seven elements south of the Jequitinhonha River. Thus, this
geographically variable character cannot be considered diagnostic using the
criteria put forward by Isler et al. (1997).
“A key issue appears to be that Stopiglia included vocal samples
from Chapada Diamantina in their analysis of cinerea and then found the
result that “Our analysis shows that the number of elements in the first
syllable of S. whitneyi ranges from zero (absent, see ‘‘Methods’’) (Boa
Nova, Bahia and Almenara, Minas Gerais) to six (Bonito and Lenc ̧o ́is, Bahia)
(Fig. 12b–h), whereas in S. ruficapilla it ranges from one (Poc ̧o
Dantas, Minas Gerais), to seven elements (Rio Novo, Minas Gerais) (see Table
3).” Worth noting that Bonito and Lençois are precisely the Chapada Diamantina
samples and are where they found a high number of introductory notes that they
(apparently) did not find in the “typical” cinerea range.
“The paper doesn’t provide details of how many notes were
associated with each sampled individual or with each sampling region, nor does
it provide any mean number of notes data, and unfortunately their recordings
are not publicly available and the email for the Elias Coelho Sound Archive
bounces.
“Regardless of how you try to read between the lines of their
analysis of vocal non-diagnosability, and with the caveat that we don’t have
access to Stopiglia’s recordings, publicly available recordings (XC and
Macaulay) paint a quite different picture and suggest that the birds are
largely readily diagnosable to one of the three vocal groups: classic ruficapilla,
classic cinerea, and classic infuscata. Of course, as Santiago
pointed out, the birds in Chapada Diamantina fit the ruficapilla vocal
pattern but Batalha-Filho’s haplotype network places them with cinerea.
“Per Van’s request here is an attempt to summarize typical
vocalizations, to relate them to Stopiglia’s terminology and findings, to
provide a couple links to typical vocalizations:
cinerea - a repeated hiccupping vocalization that consists of a softer
first note / phrase and a harder/louder second note/ phrase. At times in a lot
of the recordings it sounds like there is a bit of a “1 and a half note” first
phrase if you listen in detail:
https://xeno-canto.org/938458 (this is a good example to my ear of
being able to hear more introductory notes)
ruficapilla - a longer series of introductory notes
that also changes the timing between the final notes by quite a bit, changing
the feel of how fast the bird is vocalizing - though it is of course hard to
know what role the bird’s state of agitation plays in how often it repeats its
song
https://xeno-canto.org/385431 (one of the faster recordings I can find
where the introductory notes are more quickly stuttered, but in this case the
bird is perhaps less agitated and is not repeating the song as frequently)
Chapada Diamantina birds - basically sound
like ruficapilla
infuscata - essentially just repeats what would be
the second phrase of the vocalizations of ruficapilla/cinerea,
sometimes with 3-4 notes, sometimes in a long series - here as examples of both
the short and long series:
https://xeno-canto.org/712287 (a duet of shorter series)
https://xeno-canto.org/843883 (a long series from one bird)
The best recordings I can find from near
the zone of contact / intergradation that might represent intergradation are
these from near the Mata do Passarinho reserve:
https://ebird.org/checklist/S62644490
https://ebird.org/checklist/S52374129
https://ebird.org/checklist/S43462202
And these from a bit further north from
XC:
“I didn’t find any recordings of the birds from Vila Rica region
of Mato Grosso, but they don’t seem to be under discussion here so not really
relevant.
“With the caveat that I’m doing this as a layman, having now
listened to a few hours of audio and read the involved papers a couple times,
Stopiglia’s vocal analysis is pretty hard to extract meaningful information
from given their inclusion of Chapada Diamantina birds in cinerea and
the presentation of results that lack sufficient details about mean number of
introductory notes and about how many outlying recordings they had and from
precisely where. The neat page of clinal graphs is particularly useless in my
opinion if one wants to look at it in terms of contact zones and considering
the Chapada Diamantina birds might not just be cinerea. It would be nice
to have access to all their vocal material but based upon what is available and
field experience, I personally view the statement that “this geographically
variable character cannot be considered diagnostic” to be fairly misleading. The
fact that the vocal types and the haplotype network don’t line up neatly is
certainly interesting with respect to the Chapada Diamantina birds (and I won’t
try to get involved in the genetics discussion), but the vocal types as known
are quite diagnosable and there is a lack of available recordings from near to
the contact zone to try to examine the vocal case in that region in more
detail.”
Comments from Remsen: “NO. With
the comments from Dan and Josh, this all needs to be straightened out formally
before making a taxonomic decision.”
Comments from Areta: “I´ve been scratching my head for several
years on the Stopigila et al. (2013) paper on this complex, until Batalha-Filho
et al. (2019) appeared, and everything made more, but not complete, sense. It
is clear that there is a transition area, in which cinerea and ruficapilla
hybridize (seemingly to the south and north of the Jequitinhonha river), while
the Chapada Diamantina populations with ruficapilla voice and cinerea
mtDNA can perhaps be explained away by former reproductive contact between
them, which does not seem to have carried much into their genomes (or has it?)
or phenotypes. It is relevant that this population is so remarkably ruficapilla-like
in vocalizations, or at least that it seems to sound like birds with mtDNA
placing them in the northern ruficapilla
clade. The divergence between cinerea and ruficapilla is quite
recent (around 0.9 million years; although there is a wide posterior
probability interval: 95% highest posterior density [HPD] 0.541–3.461 mya). The
bioacoustic analyses in Stopiglia et al. (2013) could have been neater, but
nonetheless they seem to have broadly predicted the geographic patterns later
uncovered in genetic terms: there is an area of vocally intermediate birds more
or less where the genetics indicate admixing. I don´t like the lumping of all
the vocalizations in "5 degree blocks", especially because one of the
blocks seems to include samples to the north and south of the Jequitinhonha
river where one would like to see in more detail what is going on, and also
because the vocally ruficapilla-like Chapada Diamantina population has
been subsumed under the cinerea vocal dataset. Thus, I have asked Marcos
Raposo and Renata Stopiglia for a table of raw data, to see if we can refine
our understanding of what is going on here.
“However, preliminarily, the data seems
to be showing fairly stable and diagnostic vocal types that have a transition
area in which they blend into something intermediate. The question seems one of
degree here, as the zone of intermediacy is there, but it apparently has not
been thoroughly characterized at the genetic or vocal level. In the end, it
seems that the question is whether what is going on at the interaction zone is
relatively free interbreeding or conversely the populations are just hybridizing
over a narrow front. Although some nuclear gene clines seem rather wide,
Batalha-Filho et al. (2019) state that "these
analyses suggest that introgression is somewhat geographically limited, despite
hybrids being common at this locality." (p. 688; referring to locality 7
in their study), and also that "Structure results indicated a higher level
of admixture in locality 7, with an abrupt decrease in introgression levels
away from this population (Fig. 2d)" (pp. 683-684). The cline widths shown in their
Figure 4 are of course influenced by the sampling localities, and I wonder
about the nuclear composition of the Chapada Diamantina population (is it, for
the sampled genes, just like other cinerea populations?). They conclude that
there is likely selection against the hybrids, and possibly incomplete lineage
sorting as well in nuclear DNA. Thus, I would take the cline widths with a
grain of salt until there are genomic data including more nuclear genes. The
proposal states that "The narrow
cline for one Z-linked locus suggests some selection on the hybrid zone between
cinerea and ruficapilla, although there is extensive gene flow on
the rest of the genome", but such an extensive gene flow at the
genome level remains to be shown and this conflicts with the view of
Batalha-Filho et al. (2019). At any rate, one has at least to accept
that there is no full reproductive independence between cinerea and ruficapilla,
with the second question being whether there is "essential"
reproductive independence between them under a BSC framework.
“Before casting my vote,
I would like to examine the raw vocal dataset used by Stopiglia et al. (2013)
in more detail. For example, it could well be that there is clinal variation in
the number of notes in Synallaxis
ruficapilla, with an abrupt switch to the S. cinerea song through a small area in which birds sound
intermediate (in consonance with what genetic data indicates). Also, the
southern population of ruficapilla
has a distinctive inflection at the onset of the final note, which seems to be
lacking further north, and which might also exhibit clinal variation. Thus,
distinguishing several different scenarios is the task we should be up to, and
I am not sure that the available data allows us to clearly distinguish between
them. The case is difficult, given the lack of clearcut limits that are the
hallmark of classic speciation.
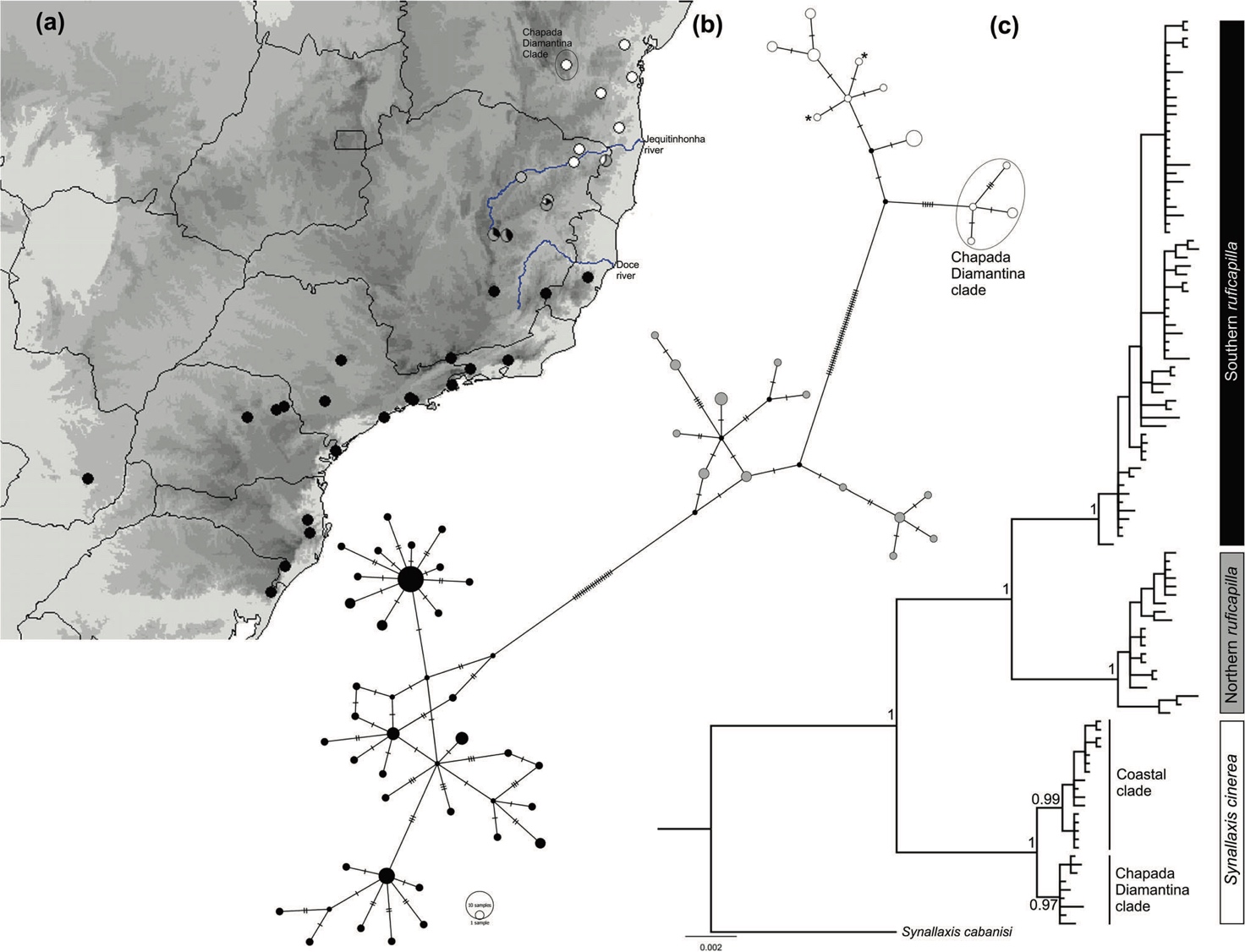
Fig 1 from Batalha-Filho et al. 2019
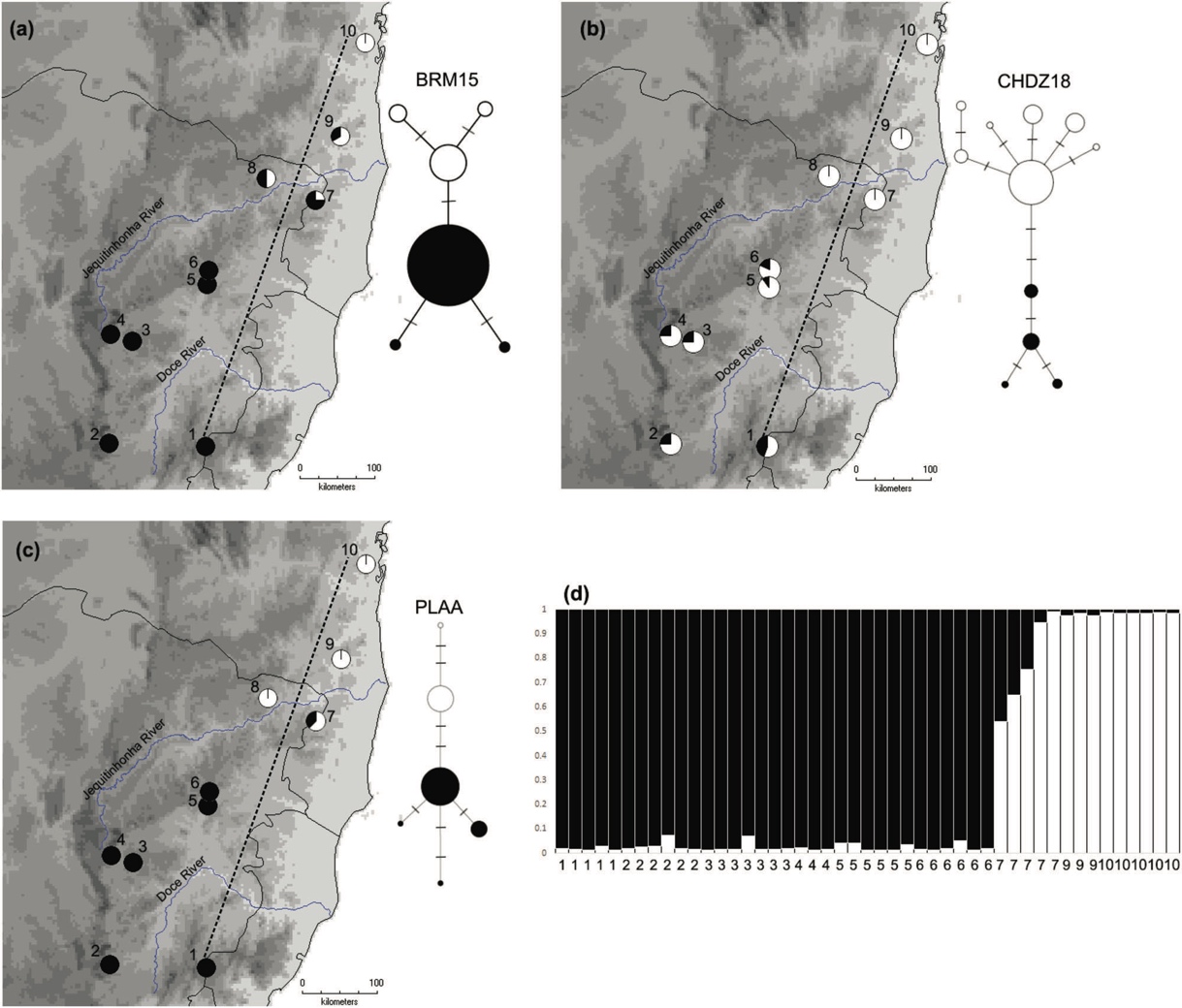
Fig 2 from
Batalha-Filho et al. 2019.”
Additional comments from Stiles: “I am
willing to change my vote to NO, based on the commentaries by Josh and Nacho.”
Comments from Areta: “NO.
Despite a lengthy email exchange, the lead and corresponding authors were
unable to provide a raw data spreadsheet. So, as things currently stand, I
cannot differentiate the different possible scenarios, and thus I prefer to
wait for more and better evidence before changing our current taxonomy.”