Proposal (1031) to South
American Classification Committee
Revise species limits
in the Schiffornis turdina complex
Background:
The
Schiffornis turdina complex currently comprises thirteen taxa sorted into
five species:
· S. turdina (5 ssp.: turdina,
intermedia, wallacii, amazonum, steinbachi)
· S. stenorhyncha (2 ssp.: stenorhyncha
and panamensis)
·
S. veraepacis (4 ssp.: veraepacis,
dumicola, acrolophites, rosenbergi)
· S. aenea (monotypic)
· S. olivacea (monotypic)
·
They
were all formerly treated as a single species (Snow
2004, Kirwan and Green 2012), although many authors have noted — based
on marked vocal differences observed in the field — that more than one species
was involved (e.g., Ridgely and Tudor 1994).
The complex has been broadly treated as consisting of five species
since ~2012 after the works of Nyári (2007) and Donegan et al. (2011). Briefly,
Nyári (2007) examined mtDNA and vocal variation in the complex, and proposed that it
should be split into five species. The proposal, however, was not widely
accepted for multiple different reasons (see Proposal 327 for details). Donegan et al. (2011) conducted an additional
analysis of vocal variation and made a new proposal with more evidence for
splits (see Proposal 505 for details). Together, the findings
of Nyári (2007) and Donegan et al. (2011) led
to the current treatment of the S. turdina complex into five species.
As
several noted in their comments on previous proposals on the S. turdina
complex, those splits were a significant step forward in recognizing
species-level diversity within the complex. However, several gaps remained, and
recognition of additional species was likely with more detailed information.
Notably, several taxa remained poorly sampled and vocal differences had never
been quantified.
New
information:
Lima et al. (2024) recently examined geographic variation in song,
plumage color, and morphology and population genetic structure across the genus
Schiffornis. They analyzed the songs of 314 individuals, 598 study
skins, 56 UCE sequences, and hundreds of mtDNA sequences, including all or
almost all taxa in each data set.
Lima et al. (2024) found eleven phenotypically and genetically
diagnosable taxa in the S. turdina complex, including a new taxon and
one taxon that has been treated a synonym of S. t. amazonum. They
corroborated previous splits and suggested that as many as nine biological
species should be recognized in the complex based on a “yardstick” approach, using patterns of
divergence between sympatric or parapatric taxa within the genus to assess the
rank of the remaining, allopatric taxa.
Patterns of phenotypic and genotypic differentiation vary widely
among taxa in the S. turdina complex. Some taxa have markedly distinct
songs but shallowly diverged genotypes; others have similar songs but deeply
diverged genotypes; others have deep mitochondrial but negligible nuclear
differentiation. Several taxa are indistinguishable visually. See Lima et al.
(2024) for details. Because variation in plumage and morphology is probably of
little (if any) relevance in defining biological species limits within this
group, I will focus below on the vocal and genetic results.
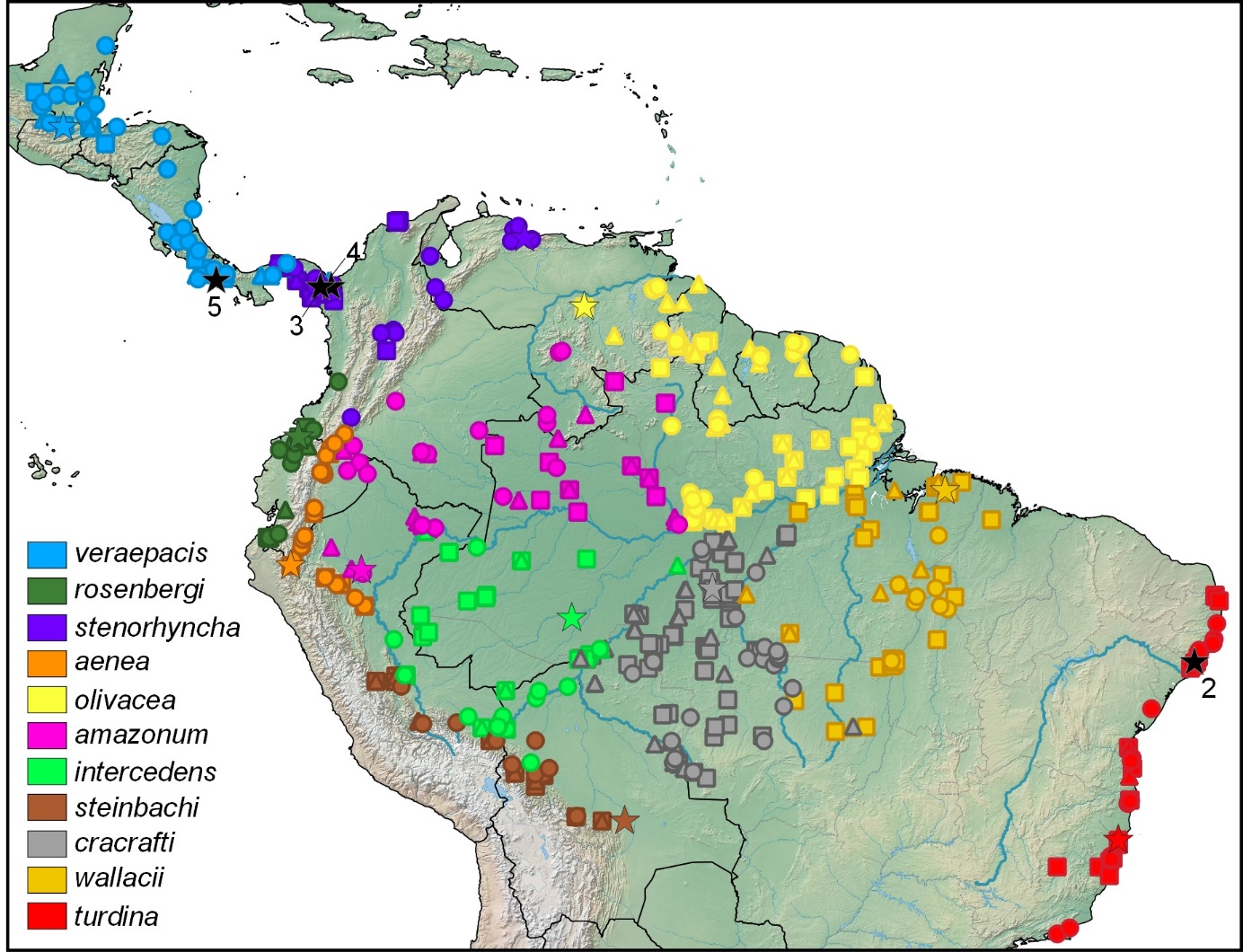
Here’s a map from Lima et al. (2024) that will be useful in
understanding the geographic range of each taxon. The map shows sampling
locations of sound recordings (circles), study skins (squares), and genetic
samples (triangles). Colored stars indicate the type locality of valid taxa
based on the revised classification proposed by Lima et al. (2024), and
numbered black stars indicate the type locality of taxa synonymized therein.
For more details and other relevant figures, see Lima et al. (2024).
The
proposal is subdivided as follows:
A.
Treat rosenbergi as a separate species from S. veraepacis
Schiffornis
veraepacis
currently comprises two disjunct populations: one in Central America (including
ssp. veraepacis, dumicola, acrolophites) and the other
ranging from western Colombia to northern Peru west of the Andes (rosenbergi).
The range limits of the four putative taxa within S. veraepacis, based
on plumage color variation, have never been fully understood. Previous work on
the S. turdina complex showed that these two disjunct populations of S.
veraepacis differ diagnosably in song and mtDNA (Nyári
2007, Donegan et al. 2011). Based on these differences, some considered
that these populations could be treated as separate species. However, some
considered that the differences between the two could reflect sampling gaps
(see Proposal 505).
Using
larger sample sizes, Lima et al. (2024) showed that
the Central American and South American populations of what is currently
recognized as S. veraepacis do have markedly different songs, with
differences equivalent in magnitude to those between the sympatric S.
veraepacis and S. stenorhyncha. Additionally, Lima et al. (2024) also confirmed, with greater
sampling, that the Central American and South American populations of S.
veraepacis are diagnosable on the basis of mtDNA. There are no consistent
differences in either phenotype or genotype within these two vocally and
genetically distinct populations. Taking all these results together, Lima et al. (2024) suggested that the Central
American and South American populations of S. veraepacis should be
recognized as two separate biological species, both monotypic: S. veraepacis
and S. rosenbergi. I recommend a YES vote on this subproposal,
primarily due to significant vocal differences distinguishing rosenbergi from
all other taxa in the complex (Lima et al. 2024).
B.
Treat steinbachi as a separate species from S. turdina
Lima et al. (2024) discovered that two vocally and genetically
distinct populations currently considered conspecific within S. turdina
are locally sympatric in northwestern Bolivia and southeastern Peru. These
populations are steinbachi and intercedens, the latter currently
considered a synonym of amazonum (more details about the name intercedens
below in subproposal “D”). These two populations differ diagnostically in
multiple song characters, and there is no phenotypic evidence of hybridization
between the two despite the close geographic proximity and even overlap of
sound recordings. Although there is some genetic evidence of hybridization
between the two, a genome-wide FST = 0.3 suggests that steinbachi
and intercedens remain considerably differentiated despite some gene
flow where they come into contact (Lima et al. 2024).
Although additional research of the contact zone between steinbachi
and intercedens is clearly needed to determine levels of reproductive
isolation, current evidence suggests that reproductive isolation between the
two is moderate at least (Lima et al. 2024). Based on the vocal differences
between the two and the sizeable genotypic differentiation in the face of some
gene flow, I believe that steinbachi and intercedens are best
considered separate biological species now.
C.
Treat amazonum as a separate species from S. turdina
What
is currently understood as S. t. amazonum actually includes two vocally
and genetically distinct populations separated by the Amazon River (more
details below in subproposal “D”). Here in subproposal “C” we are dealing with
the northern population. This population exhibits significant vocal and
genome-wide differentiation in relation to all other taxa of the S. turdina complex
except the newly described cracrafti (Lima et
al. 2024).
See
subproposals “F” and “I” before voting on this one.
D.
Treat intercedens as a separate species from S. turdina
Schiffornis
turdina intercedens
Todd, 1928 was synonymized with S. t. amazonum (Sclater, 1860) shortly
after its description (Hellmayr 1929) and has
since been universally treated as a synonym (Pinto
1944, Snow 2004, Kirwan and Green 2012). Using the largest sample of
specimens examined to date, Lima et al. (2024)
corroborated Hellmayr’s (1929) assessment that the two taxa are indeed
visually indistinguishable. However, Lima et al.
(2024) found that intercedens and amazonum have
diagnostically distinct songs and genotypes, and therefore reinstated intercedens
to taxonomically recognize these differences.
This
population, intercedens, exhibits significant vocal and genome-wide
differentiation in relation to all other taxa in the S. turdina complex
except wallacii and cracrafti. It differs diagnostically from
both in song and mtDNA, but vocal differences between intercedens and wallacii
are subtle and — based on a “yardstick” approach — unlikely to generate
premating isolation (Lima et al. 2024).
See
subproposals “E” and “H” before voting on this one.
E.
Treat wallacii as a separate species from S. turdina
This
taxon caused some confusion in previous proposals on the S. turdina
complex owing to uncertainties regarding its type locality. Many reference
works incorrectly mention that the range of wallacii reaches the Guiana
Shield (e.g., Hellmayr 1929, Pinto 1944, Kirwan and Green 2012), where only olivacea
occurs (Lima et al. 2024). Given that wallacii and olivacea are
visually indistinguishable (Lima et al. 2024) and their ranges were
traditionally based on plumage color variation alone, this confusion is not
surprising. Donegan et al. (2011) correctly assigned the type locality of wallacii
to the city of Belém, in the Brazilian state of Pará. Lima
et al. (2024) further restricted the type locality to the vicinities of the
Guamá River near Belém, based on the collector’s itinerary.
Lima et al. (2024) found sizeable vocal and genome-wide differentiation
between wallacii and all other taxa in the S. turdina complex
except cracrafti and intercedens. Although wallacii
differs diagnostically from cracrafti and intercedens in song and
mtDNA, vocal differences between wallacii and intercedens are
subtle and — based on a “yardstick” approach — unlikely to generate significant
reproductive isolation (Lima et al. 2024). Song differences between wallacii
and cracrafti are equivalent in magnitude to those between sympatric
Schiffornis species, but overall nuclear genetic differentiation is
negligible (genome-wide FST ~ 0).
See
subproposals “D” and “H” before voting on this one.
F.
Recognize the newly described Schiffornis cracrafti as a species
Lima et al. (2024) discovered one vocally and genetically
diagnosable population largely restricted to the Madeira/Tapajós interfluvium
in southwestern Amazonia. They described a new taxon, named cracrafti,
to taxonomically recognize this population. Using a “yardstick” approach, Lima et al. (2024) inferred that cracrafti may have
evolved significant reproductive barriers from all other taxa in the complex
except amazonum.
See
subproposals “C” and “I” before voting on this one.
G.
Treat nominate turdina as a separate species from all the rest
Schiffornis
t. turdina
of the Atlantic rainforest of eastern Brazil was poorly sampled in previous
studies on the S. turdina complex (Hellmayr
1929, Nyári 2007, Donegan et al. 2011). Using a geographically
comprehensive samples of this taxon, Lima et al.
(2024) found that it differs diagnostically from all other taxa in the complex
by multiple song characters and exhibits substantial genome-wide
differentiation. It is also the most differentiated in morphology (Lima et al. 2024), though this is likely of little
relevance here. Because the levels of vocal and genetic divergence
between turdina and the remaining taxa
in the complex are equivalent to or greater than those between sympatric Schiffornis
species, this split must be straightforward.
If
subproposals A–G pass, we will not have to make combinations of taxa into
polytypic species. But if some of those subproposals don’t pass, then we will
have to deal with combinations. Here are subproposals that deal with the latter
possibility:
H.
Treat wallacii and intercedens together as a polytypic species
separate from all the rest
There
is no evidence for reproductive isolation between wallacii and intercedens
(Lima et al. 2024). Although they differ
diagnostically in song, differences are smaller than those between sympatric Schiffornis
species. Therefore, we can’t assume that they would mate assortatively based on
song differences if they were to come into contact. Additionally, they have shallow nuclear differentiation (genome-wide FST =
0.12). Therefore, we can’t assume that they would exhibit significant
postmating reproductive barriers such as genetic incompatibilities. They have
sizeable mtDNA divergence (uncorrected pairwise distance for cyt-b, ND2, and
COI: 3.7–4.4%). Is that meaningful in defining biological species limits?
Probably not.
Note
that wallacii and intercedens are not sister taxa. A NO vote on
this subproposal may therefore result in a nonmonophyletic species taxon, and
with a strange geographic distribution. If this is considered problematic, you
should vote NO on this subproposal and YES on subproposals “D” and “E.”
Otherwise, vote YES on this subproposal and NO on subproposals “D” and “E.”
I.
Treat cracrafti and amazonum together as a polytypic species
separate from all the rest
There
is no evidence for reproductive isolation between cracrafti and amazonum
(Lima et al. 2024). Although they differ
diagnostically in song, differences are smaller than those between sympatric Schiffornis
species. Therefore, we can’t assume that they would mate assortatively based on
song differences if they were to come into contact. Additionally, they have shallow nuclear differentiation (genome-wide FST =
0.16). Therefore, we can’t assume that they would exhibit significant
postmating reproductive barriers such as genetic incompatibilities. Their mtDNA
divergence, although sizeable (3.5–5%), is probably not meaningful in defining
biological species limits.
Note
that cracrafti and amazonum are not sister taxa. A NO vote on
this subproposal may therefore result in a nonmonophyletic species taxon, and
with a strange geographic distribution. If this is considered problematic, you
should vote NO on this subproposal and YES on subproposals “C” and “F.”
Otherwise, vote YES on this subproposal and NO on subproposals “C” and “F.”
References
Donegan, T. M., A.
Quevedo, M. McMullan, and P. Salaman (2011). Revision of the status of bird
species occurring or reported in Colombia 2011. Conservación Colombiana
15:4–21.
Hellmayr, C. E. (1929).
Catalogue of birds of the Americas and the adjacent islands in Field Museum of
Natural History. Volume XIII, Part VI. Field Museum Press.
Kirwan, G. M., and G.
Green (2012). Cotingas and manakins. Christopher Helm.
Lima, R. D., F.
Bocalini, and L. F. Silveira (2024). Integrative revision of species limits in
the genus Schiffornis (Aves: Tityridae) reveals cryptic diversity in the
Neotropics. Zoological Journal of the Linnean Society 200:1048–1079.
Nyári, Á. S. (2007).
Phylogeographic patterns, molecular and vocal differentiation, and species
limits in Schiffornis turdina (Aves). Molecular Phylogenetics and
Evolution 44:154–164.
Pinto, O. M. O. (1944).
Catálogo das aves do Brasil e lista dos exemplares na
coleção do Departamento de Zoologia. 2° parte, ordem Passeriformes
(continuação): superfamília Tyrannoidea e subordem Passeres. Departamento de
Zoologia.
Ridgely, R. S., and G.
Tudor (1994). The birds of South America. Volume 2 (The Suboscine Passerines).
University of Texas Press.
Snow, D. W. (2004).
Family Pipridae (Manakins). In Handbook of the Birds of the World (J. del Hoyo,
A. Elliot and D. A. Christie, Editors). Lynx Edicions, pp. 110–169.
Rafael D.
Lima, August 2024
Note from Remsen: If the
proposal passes, we will need a separate proposal on English names.
_________________________________________________________________________________________________________
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments from Remsen: “This is
an excellent compilation and synthesis of everything available on the turdina
complex. What a fascinating, complex
system, and a good example of conservative evolution in plumage.
“A. YES. Vocal differences between the two warrant treatment as
separate species. Differences in mtDNA,
however, are irrelevant to species limits in allotaxa, and I think all genetic
results that conflict with other data in this analysis should be treated with
caution – they are mtDNA gene trees that do not necessarily reflect historical
branching patterns.
“B. YES. Vocal differences combined with no evidence of free gene
flow despite parapatry or sympatry essentially require these two to be treated
as separate species.
“C. YES to be consistent in using voice as an indicator of species
limits.
“D. NO based on lack of strong vocal differences.
“E. YES based on vocal differences.
“F. YES based on vocal differences.
“G. YES. Easiest decision
of all.
“H. YES. Tough one! It
looks like the intervening taxon cracrafti has diverged from the
ancestral form more rapidly than intercedens and wallacii have,
leaving them with a bunch of shared ancestral vocal characters, which poses
problems for taxonomy. So, the BSC
classification here is uncomfortable if intercedens and wallacii (which
has priority) are treated as conspecific.
I think that’s better than treating the two as separate species when
they don’t differ in voice, the character most important for speciation in this
group.
“I. NO. Difficult case, and a good example of how our typological
classification scheme has difficulty with mosaic evolution. The conclusion that they are not sister taxa
based on mt DNA gene trees is premature in my opinion.”
Comments from Lane:
“A) A weak YES considering S. rosenbergi separate from S.
veraepacis. To my ear, the voices are not strongly differentiated, but I
will accept Lima et al.’s conclusions on this.
“B) YES to treating S. steinbachi as a separate species. I
think the vocal differences are quite clear to me and worthy of species
recognition.
“C) YES to considering S. amazonum separate from S.
turdina. Again, biogeographically and vocally, this is a strong case.
“D) I agree to recognizing the taxon intercedens as
separate from amazonum, but not as a separate species, so NO, I guess.
“E) This is weirdly worded. I support separating wallacii as
separate from turdina, but not from amazonum. So YES to separating it from
turdina, but NO to recognizing it as a species on its own.
“F) Similarly to the last, I say YES to recognizing cracrafti
as a taxon and as a species apart from S. turdina, but NO to recognizing
it as a species on its own. The evidence suggests to me that cracrafti
is best considered part of a polytypic S. amazonum.
“G) YES. Vocally, molecularly, and phenotypically, I think the
case is good for Atlantic Forest S. turdina being separated from the
rest of the complex as a biological species, even if doing so results in a
non-monophyletic S. amazonum.
“H) NO. In Lima et al (2024) the branching phylogeny published is
a cytB-based tree, and I am reluctant to accept that branching scheme at face
value (as Van seems reluctant to do). I would prefer using audio characters as
my best guess to taxonomic and species-level affiliation, and to my ear these
Amazonian forms are all quite similar (and in my experience, motivation, such
as response to playback, can affect song structure enough that I would be
reluctant to use anything short of a large sample of unsolicited song
recordings to determine vocal characters). Thus, I would accept a polytypic S.
amazonum that encompasses cracrafti, wallacii, intercedens,
but apart from S. turdina (even at the risk of a non-monophyletic S.
amazonum), which is not an option in the present set of votes.
“I) NO. See H.”
Additional comments from Rafael
Lima: “"I personally have no objection to nonmonophyletic species
taxa and am comfortable to any lumping of Amazonian populations, given that
evidence for reproductive isolation is equivocal or weak for several of them.
However, I want to clarify that the phylogenetic relationships mentioned in
both the paper and this proposal are not based solely on mitochondrial markers.
Lima et al. (2024) conducted several different phylogenetic analyses using both
mitochondrial and genome-wide nuclear markers (specifically UCEs), and these
analyses were largely congruent. When the proposal mentions that certain lumps
may result in nonmonophyletic species taxa, this includes nonmonophyly across
all phylogenetic analyses performed. For example, the conclusion that cracrafti
and amazonum are not sister taxa is based on concatenated and coalescent
analyses of UCEs as well as mtDNA data. Also, a polytypic S. amazonum
encompassing cracrafti, wallacii, and intercedens, but
excluding S. turdina, as Lane suggests, would form a nonmonophyletic
taxon based on genome-wide markers (see Supplementary Figures S13–15 in Lima et
al. 2024). I am comfortable with such a taxon, though."
Comments
from Zimmer:
“
“A)
YES. Based upon vocal differences. Also, it strikes me as an odd distribution
that rosenbergi would be conspecific with polytypic populations in
Central America (ssp. veraepacis, dumicola, acrolophites) that are
separated from an isolated, monotypic rosenbergi in South America by the
range of a different species (S. stenorhyncha), that is sympatric with veraepacis.
“B) YES to
treating steinbachi as distinct both from S. turdina, and, as
distinct from intercedens, based on differences in multiple song
characters and lack of evidence of free gene flow between steinbachi and
intercedens despite the two existing either in parapatry, or possibly
sympatry.
“C)
YES. Based on vocal differences and
biogeographic considerations, this seems straightforward.
“D) This is
where I’m starting to get confused as to what I am actually voting on, and, in
looking at Van’s and Dan’s comments, it looks like we are all interpreting the
question differently. This subproposal
reads: “Treat intercedens as a
separate species from S. turdina.”
Taken at face value, I would vote YES on recognizing intercedens
as being distinct from nominate turdina (See Subproposal G – which is to
treat turdina as distinct from everything else.), based upon
“significant vocal and genome-wide differentiation in relation to all other
taxa in the complex except wallacii and cracrafti.” (Lima, in the
Proposal). So, my YES vote is only to
treat intercedens as separate from nominate turdina, not as
separate from wallacii or cracrafti. Implicit to me was recognition/reinstatement
of intercedens as a valid taxon at some level (Lima et al. 2024), rather
than retaining it as a junior synonym of amazonum. However, I see that Van has voted “No”,
citing “lack of strong vocal differences”, which would be the case if we were
considering the question of recognizing intercedens as a species
distinct from wallacii and cracrafti, but not if the question is
one of whether to treat intercedens as distinct from turdina,
which is how Subproposal D is worded.
Meanwhile, Dan agreed with recognizing intercedens as a taxon
(presumably as a subspecies) distinct from amazonum, but not “as a
species on its own” -- in other words, I
am interpreting Dan’s comments to mean that he agrees that intercedens
is a valid taxon, but that he doesn’t consider it specifically distinct from amazonum. But, again, the wording of Subproposal D is
asking whether we should consider intercedens as distinct from turdina,
not from amazonum. So, Van and
Dan seem to be basing their NO votes on the latter question, whereas my YES
vote is based upon the former question.
Dan says as much in his responses to both Part E & F.
“E) As in
my comments on Part D, I vote YES to treating wallacii as specifically
separate from S. turdina, but NO to treating it as distinct from intercedens,
cracrafti, and amazonum, based upon minimal vocal differences. I would vote YES to treating these 4
Amazonian taxa as part of a polytypic S. amazonum, that is distinct from
all other members of the complex, but that doesn’t seem to be one of the voting
options in this Proposal.
“F) As in
my comments on Parts D & E, I vote YES to treating cracrafti as
specifically separate from S. turdina, but NO to treating it as distinct
from intercedens, wallacii, and amazonum, based upon minimal
vocal differences. I would vote YES to
treating these 4 Amazonian taxa as part of a polytypic S. amazonum, that
is distinct from all other members of the complex, but that doesn’t seem to be
one of the voting options in this Proposal.
“G)
YES. This one seems particularly
straightforward, on the basis of differences in multiple song characters,
genetic differences, morphological differences (although, I agree, that
morphology is probably not overly relevant in this complex), and from the
standpoint of biogeography.
“H)
NO. According to the Proposal’s summary
of Lima et al. 2024, intercedens “exhibits significant vocal and
genome-wide differentiation in relation to all other taxa in the S. turdina
complex except wallacii and cracrafti.” And “Lima et al. (2024) found sizeable vocal
and genome-wide differentiation between wallacii and all other taxa
in the S. turdina complex except cracrafti and intercedens.” So, there are no sizeable vocal differences
between these 3 taxa, and therefore, I don’t see the basis, other than the
genetic data, for treating wallacii and intercedens together as a
species that does not include cracrafti and amazonum.
“I) NO.
According to the Proposal’s summary of Lima et al. 2024, “There is no evidence
for reproductive isolation between cracrafti and amazonum(Lima et
al. 2024). Although they differ diagnostically in song, differences are smaller
than those between sympatric Schiffornis species.” So, if the vocal differences between cracrafti
and wallacii & intercedens are unremarkable, and there is no
evidence for reproductive isolation between cracrafti and amazonum,
which also do not differ strongly from one another vocally, then I see no
strong reason for separating the latter two taxa from wallacii + intercedens. Again, my YES vote would be for a polytypic S.
amazonum (amazonum + cracrafti + intercedens + wallacii) as separate
from everything else and based upon lack of yardstick-level vocal
differentiation between the 4 taxa involved.”
Comments from Areta: “I´ve been struggling to integrate the
vocal and genetic (mtDNA and UCE datasets) in order to have a firmer grasp of
the taxonomy of these Schiffornises. My final take is like that of Kevin and
Dan. Although I am conflicted by the non-monophyly of the polytypic S. amazonus (including intercedens, cracrafti, and wallaci),
and by the "leapfrog" pattern of taxa with 3 (amazonus and cracrafti)
and 4 (intercedens and wallaci) notes, there are enough
uncertainties in the phylogenetic relationships suggested by the different DNA
sources to cast doubt on what the true relationships are here. Keeping a
polytypic amazonus seems the
conservative stance, and while the vocal differences seem consistent, it is not
exactly clear what do they mean from the perspective of interbreeding, given
also how similar these vocalizations sound.
“I
am also somewhat conflicted by the split of the sister, little diverged, and
vocally similar rosenbergi from veraepacis, mostly because of the
apparently fairly widespread vocal evidence of hybridization in Panama between
the phylogenetically distant and vocally divergent veraepacis and stenorhyncha.
I wonder whether these vocally intermediate individuals are really hybrids.
“It
seems to me that things are more fluid in Schiffornis
than what discrete categories might be able to describe, and further genomic
work will surely come up with cool and taxonomically useful information.”
“A. YES, but quite
conflicted on this one: the more I think on this, the less convinced I am of
splitting veraepacis from rosenbergi (see comments above)
B. YES
C. YES, but see D, E,
F, H, and I to understand what should be included in S. amazonum (and
see comments above; no solution pleases me fully).
D. YES to treating intercedens
as separate from turdina, but no to S. intercedens as a different
species level taxon. Treat intercedens as part of a polytypic amazonum.
E. YES to treating wallacii
as separate from turdina, but no to S. wallacii as a different
species level taxon. Treat wallacii as part of a polytypic amazonum.
F. YES to treating cracrafti
as separate from turdina, but no to S. cracrafti as a different
species level taxon. Treat cracrafti as part of a polytypic amazonum.
G. YES to a monotypic S.
turdina.
H. NO, treat wallacii,
intercedens, cracrafti, and amazonum as part of a
polytypic amazonum.
I. NO, treat wallacii,
intercedens, cracrafti, and amazonum as part of a
polytypic amazonum.”
Comments from Stiles: “Schiffornis
splits: the problem here is that the wording is confusing or too vague in some
parts.
A: split rosenbergi
from veraepacis- YES
B. split steinbachi
as a species from turdinus - YES (interesting that this species takes
off as a foothill species south of where (after a gap) aenea left off.
C. split amazonum
from turdinus as a species (?):YES. Here starts the paring off of
various taxa from turdinus.
D.
recognize taxon intercedens and split from turdinus.
E. ditto
for cracrafti
F. ditto
for wallacei (although suggested that all should be called species, this
seems to have confused some because all were later demoted to subspecies;
perhaps here simply name them all as taxa.
G. Split turdina
from everyone else (as it has been left monotypic).
H: combine wallacii
and intercedens as a species, and
I: ditto cracrafti
and amazonum - both based on similarity of vocalizations but both
conflicting with (mtDNA) phylogeny and (at least in part) with distributions.
More doubts - but the beginning of a possible consensus? Dan, Kevin and Nacho
have suggested that all taxa in H and I
be combined in a single polytypic species (amazonum has priority). This
also seems to me the best solution, This was not presented as an option in the
original proposal, but it could easily be accommodated as option J (which also
appears be acceptable to Rafael).
Comments from Robbins: “I’ve gone through this proposal twice,
about a month apart. Based on the vocal
and limited genetic data (we know the issues associated with mitochondrial
data), it isn’t clear what is the best course of action for some of these
subproposals. However, in the end, I too
agree with what Kevin and Dan have concluded. Thus, I vote as follows,
primarily based on what has already been stated.
“A. A “lukewarm”
endorsement of voting YES, as the vocalizations are quite similar.
B. YES, the vocal
differences are clearly different.
C. YES, this is one of
the more clearcut cases.
D. Kevin makes a good
point of what are we actually voting on in this proposal. I agree with him, if
the question is treating intercedens separate from turdina, then
I vote YES. But, look how I vote in E & F.
E. YES. Kevin has
clearly laid out my thoughts on this.
F. YES. Ditto to what
Kevin states.
G. YES, song and
genetic data makes this, like B & C, a straightforward decision.
H. NO for reasons that
have been stated by other members.
I. NO for reasons that
have been stated by other members.”
Comments from Naka: “This case is indeed
quite complex. I have spent over a week thinking how to best solve the
taxonomic mess. I have gone from many yeas to many nays in my votes.
Unfortunately, voice, DNA, and distribution do not seem to tell a clear story,
and morphology isn’t offering many tips. Therefore, I am having a tough time
having to decide whether DNA or voice or distribution makes more sense in this
case. I commend the authors of the proposal for presenting such a rich
dataset in their article (+ Supp. material), but the data presented in this
proposal is limited to fully understand the case. I wish trees, structure
plots, and spectrograms would have been included here, as I had to go back and
forth many times. Unfortunately, I think the wording of the proposals and
subproposals is not ideal and somewhat confusing at times.
“Despite the wording of the proposal, I think
that for the main problem (S. amazonum and turdina) we have four
alternative paths. I have included all four below, which I ordered by my own
increasing preference. I think path 1 is worse than 2, 2 worse than 3, and 3
worse than 4.
“I will offer my votes, which are consistent
with my view that the path 4 (see below) is the most consistent and less
problematic.
“Path 1. Virtually every vote cast seems to be
happy creating a paraphyletic clade, where a polytypic S. amazonum
includes cracrafti + intercedens + wallacii, excluding S.
turdina, as a monotypic taxon. The problem I see with this proposal is that
not only champions a paraphyletic taxon, but by doing so, it creates an
Amazonian Frankenstein in terms of vocalizations. I would have no problem in
having a non-monophyletic lineage excluding turdina, IF turdina
would have been an outlier of some sort. But it happens that not only wallacii
seems to be its closest relative (both in mt and nDNA), and share high
admixture in the nuclear genome, but also has the same vocal type with four
notes and relatively similar shaped notes (see types C and E in Lima's Figure
3).
“Path 2. Another option is to accept an
Amazonian/Atlantic Forest monophyletic clade with considerable vocal variation.
This taxon would include turdina + wallacii + cracrafti + amazonum +
intercedens + steinbachi within an expanded polytypic S. turdina. I
don’t think this is ideal but looks better to me than path 1.
“Path 3. We could also accept two monophyletic
clades, where S. turdina includes wallacii + cracrafti,
and S. amazonum includes intercedens + steinbachi. The problem is
that turdina's vocal type with four notes is shared by turdina,
wallacii, and intercedens, but not cracrafti. Therefore, the
leapfrog pattern mentioned by some in their votes.
“Path 4. Finally, I see an option that does not
require crossing any red lines. This includes a) having a polytypic S.
turdina + wallacii (which by the way are sister taxa, highly
admixed, and similar sounding); b) a new species cracrafti, which is a
bit problematic because it sounds a lot like amazonum; and c) a
monophyletic S. amazonum with intercedens + steinbachi.
“This
last treatment may solve the taxonomy without creating major disruptions.
“According
to this treatment, I provide my votes below:
“A. Treat rosenbergi
as a separate species from S. veraepacis: NO. The two taxa (rosenbergi
and veraepacis) are each other's closest relatives and, according to
Lima's Figure 3 do not seem to show such a strong vocal variation, I am happy
to go with two allopatric populations, shared by the Chocó and C. America.
This would not be the first such distribution, and I think is quite interesting
and educative in terms of understanding these two regions’ biogeography. I
admit that the presence of steinbachi makes this story quite
challenging, though. However, I don’t know any of these taxa, so I am happy to
change this vote under pressure.
“B. Treat steinbachi
as a separate species from S. turdina: YES, I am happy to treat steinbachi
as a separate species from S. turdina, but I don’t think this is the
point here. I agree that steinbachi should be included within polytypic
S. amazonum.
“C. Treat amazonum
as a separate species from S. turdina: YES, I am happy to consider them
distinct species, as long as wallacii and potentially cracrafti
fall within its closest relative: S. turdina.
“D. Treat intercedens
as a separate species from S. turdina: YES, despite its vocal
similarity, I think intercedens should be split form S. turdina
but included within S. amazonum.
“E. Treat wallacii
as a separate species from S. turdina: NO. In Lima’s mtDNA tree (Figure
8), wallacii forms a well-supported clade with turdina and cracrafti,
and this close relationship remains close in the SNPs PCA (Fig. 9). Vocally
speaking, the songs of wallacii and turdina seem quite close,
with four notes and similar shapes. Biogeographically, other groups show the
same close relationship between Easternmost Amazonian populations (Belem area)
and the Atlantic Forest taxa, which seems to be the case here. Quite interestingly,
Limas’s figure 9, shows a very high admixed populations between the Atlantic
Forest samples and those from Belem.
“F. Recognize the newly
described Schiffornis cracrafti as a species: A tentative YES. I don’t
like the idea of keeping this taxon with closely related, yet different
sounding turdina + wallacii, but at the same time, its voice
looks surprisingly similar to that of S. amazonum.
“G. Treat nominate turdina
as a separate species from all the rest: NO. As I mentioned before, I think
this taxon should include at least wallacii, from the Belem Area of
endemism.
“H. Treat wallacii
and intercedens together as a polytypic species separate from all the
rest: NO. I don’t like the idea of creating this paraphyletic taxon, solely
based on song and excluding the genetic data available.
“I. Treat cracrafti
and amazonum together as a polytypic species separate from all the rest:
NO. I don’t like the idea of creating this paraphyletic taxon, solely based on
song and excluding the genetic data available.”
Additional
comments from Rafael Lima: “First, I’d like to
apologize for the confusing wording of the proposal. At the time, it seemed
that the most concise way to present the many taxonomic alternatives was
through a dichotomous and sequential arrangement of subproposals. However, I
now realize that I did a poor job of explaining how this structure was meant to
be interpreted. My intention was that once a particular subproposal receives a
YES vote, the corresponding taxon would be considered resolved and excluded
from all subsequent subproposals. I also assumed that everyone voting would
read the full paper before casting a vote, so the proposal itself only needed
to summarize the evidence briefly. After all, this checklist is a formal
publication, and the paper will be cited to support its conclusions, so I
assumed the authors of the checklist would read their references thoroughly. In
any case, despite the initial lack of clarity, we appear to be getting closer
to a resolution.
“The situation with Schiffornis
is indeed complex, and any attempt to impose a categorical scheme will
inevitably be controversial. That’s why, in the paper, we prioritized
discussing the conflicts among different lines of evidence rather than offering
overly simplified taxonomic conclusions. It should be clear that this kind of
“messiness” is unlikely to be resolved by new data in the future; the nature of
variation in this group will probably pose a persistent challenge for taxonomy.
In fact, as more taxonomic studies integrate multiple phenotypic traits and
genetic markers, cases of discordant patterns like this one will likely become
increasingly common.
“That said, we still need to
categorize these lineages into species taxa. The ‘best’ solution obviously
depends on how one defines ‘species.’ In this context, Naka’s proposal gives
priority to phylogenetic relationships over reproductive isolation; it aims to
avoid nonmonophyletic species, even if that means recognizing taxa with weak
reproductive isolation as distinct species. My understanding is that this
approach is not consistent with the committee’s stated use of the BSC. For
example, I don’t see how cracrafti could be considered a distinct
biological species from amazonum. The fact that they are not sister taxa
is irrelevant under the BSC.
In my view, the
classification that best reflects the current evidence and aligns with the BSC
is as follows:
- Schiffornis turdina
- Schiffornis amazonum (including cracrafti as a subspecies)
- Schiffornis intercedens (including wallacii as a subspecies)
- Schiffornis steinbachi
- Schiffornis stenorhyncha
- Schiffornis veraepacis
- Schiffornis rosenbergi
- Schiffornis aenea
- Schiffornis olivacea
“Let’s call this ‘Option A.’ This
classification considers only evidence of reproductive isolation, as I believe
any BSC-based classification should. Monophyly at the species-subspecies level
is not the primary criterion.
“Another possibility, raised
by several members here, would be:
- Schiffornis turdina
- Schiffornis amazonum (including intercedens, wallacii,
and cracrafti as subspecies)
- Schiffornis steinbachi
- Schiffornis stenorhyncha
- Schiffornis veraepacis
- Schiffornis rosenbergi
- Schiffornis aenea
- Schiffornis olivacea
Let’s call this “Option B.”
I’m also fine with this one. I have no strong preference between A and B, given
that the evidence for reproductive isolation among the Amazonian taxa is
equivocal.
“A third possibility would
be:
- Schiffornis turdina (including amazonum, intercedens, wallacii,
and cracrafti as subspecies)
- Schiffornis steinbachi
- Schiffornis stenorhyncha
- Schiffornis veraepacis
- Schiffornis rosenbergi
- Schiffornis aenea
- Schiffornis olivacea
“This would be “Option C.”
I’m less comfortable with this alternative because turdina is strongly
differentiated from the Amazonian taxa in the nuclear genome, suggesting that
significant postzygotic barriers are plausible.
“I hope this summary
clarifies the options. I would be comfortable with any of them, as all are
supported by the available evidence and ultimately come down to one's
preferences. That said, I do believe Options A and B are preferable to Option
C.”
Comments from Claramunt: “A
complex case. Levels of lineage sorting are very high in the mtDNA (as usual),
suggesting a strong geographic structure and a specific history for the
complex. Maintaining mtDNA exclusivity despite presumed broad areas of
parapatry along the E Andean foothills suggests intrinsic reproductive
isolation between the foothill taxon aenea and the Amazon lowland taxon
amazonus.
“The phylogenetic analysis of UCEs
seems to confirm several aspects of the mtDNA topology, like the basal position
of Guianan and a N Andes-Central American clade (aenea, stenorhyncha,
veraepacis). Also notable are the turdina samples that are nested
withing wallacii, suggesting that the Atlantic forest form is derived
from an E Amazonia stock. However, the UCE population structure analysis (Fig.
9), suggests gene flow and mixed ancestry across Amazonian and Atlantic forest
forms, including the S Andes foothill steinbachi. Four ancestral
populations are inferred but there are many specimens of admixed ancestry, and
the pattern looks more like isolation by distance, not reproductive isolation
or separate lineages. I’m not convinced by the argument that there may be
post-zygotic genetic incompatibilities just because the genomes have
accumulated some differences. But it is possible that the pop. structure
analysis is flawed. There are probably too few specimens for some clades for
the algorithm to claim a distinct ancestral population. For example, olivacea
looks like a divergent taxon in both mtDNA and UCE trees but had only two
samples in the structure analysis. For that reason, I don’t think we have a
good picture of what’s going on at the genomic level. At face value, the
structure analysis suggest we are dealing with a single species with some
ancient genetic structure but free gene flow.
“Regarding plumage and songs, the
pattern seems like two different incongruent patchworks. I don’t think a
yardstick for inferring reproductive isolation is sound, especially in cases
like this in which it is based on very few sympatric pairs, and then extrapolated
to the entire complex.
“So, regarding the specific
proposed changes:
A. Treat rosenbergi
as a separate species from S. veraepacis. YES. Reciprocally
monophyletic mtDNA and they seem to differ in plumage and voice in a diagnostic
way (Fig. 6). They show a very shallow divergence and there is a big sampling
gap in the Chocó, but I give them the benefit of the doubt.
B. Treat steinbachi
as a separate species from S. turdina. NO. turdina is
nested within the Amazonian complex -- it is not a separate lineage.
C. Treat amazonum
as a separate species from S. turdina. NO. turdina is
nested withing the Amazonian complex -- it is not a separate lineage.
D. Treat intercedens
as a separate species from S. turdina. NO. turdina is
nested withing the Amazonian complex, it is not a separate lineage.
E. Treat wallacii
as a separate species from S. turdina. NO. MtDNA is nearly
reciprocally monophyletic, but the UCE tree suggests incomplete separation: turdina
lineages are nested withing wallacii. Songs look very similar but seem
to differ in the multivariate space (Figure 4) and have diagnosable differences
in two traits. The genomic structure analysis suggests gene flow between the
two, in particular wallacii specimens show mixed ancestry between the
rest of the Amazon and turdina. It
seems like turdina was derived from an Amazonian stock and
differentiated in the Atlantic forest, being derived directly and more similar
to the closest geographical wallacii.
F.
Recognize the newly described Schiffornis cracrafti as a species. NO. it is
part of an Amazonian complex that needs more clarification.
G. Treat
nominate turdina as a separate species from all the rest. NO.
According to the mtDNA tree and the UCE topology, turdina seems to
derive from wallacii. The genomic structure analysis further suggest
gene flow (mixed ancestry) involving wallacii.
H. to treat
wallacii and intercedens together as a polytypic species separate
from all the rest. NO. The
lack of diagnostic differences may be due to retention of ancestral
morphologies. Their similarity does not indicate close affinities. Both mtDNA
and UCEs show that they are not closest relatives.
I. Treat cracrafti
and amazonum together as a polytypic species separate from all the rest.
NO. The lack of diagnostic differences may be due to retention of
ancestral morphologies. Their similarity does not indicate close affinities.
Both mtDNA and UCEs show that they are not closest relatives.
“Instead, I would suggest we could
discuss these more basic taxonomic changes:
1)
Separation the Andes-trans-Andes clade from the Amazonian group.
2)
Separation olivacea from the rest.
3) Separate
turdina from the Amazonian group.”
Comments from Stiles: “A. YES;
B.YES;C. NO; D. NO; E. NO; F. NO; G. YES, H. YES, I. YES.
Comments from Mario Cohn-Haft
(voting for Bonaccorso): “NO to all proposed changes. Do I feel
lame and on shaky ground about this? Yes. My basic argument is that the results
presented by this very important paper that makes the greatest contribution to
the subject in a long time are sufficiently unusual and difficult to explain
from an evolutionary standpoint that we should wait until the inevitable
follow-up studies give a clearer indication of what the mechanisms are to cause
such surprising and unusual results. By unusual, I’m referring to sister taxa
that are not close geographical neighbors, niche/habitat shifts that seem to
pop up inexplicably, vocal variations that don’t follow an intuitive pattern of
differentiation (or that maybe I just didn’t understand correctly), and
datasets that conflict in their conclusions. I’ll try to elaborate.
"The interplay between
vicariance or niche shift brought about by historical landscape changes versus
dispersal are expected to lead to complex patterns of speciation, anywhere in
the world. What has distinguished Amazonian result, specifically, and those
from Neotropical phylogenetic reconstruction in general is that certain
patterns repeat so frequently that they’ve established a notion of “normal”.
The results found here are far enough from normal that to believe they are
correct (and not caused by sampling or analysis issues or by processes still in
course that haven’t actually yet led to solid speciation) I would like to see
more.
“The conclusion that morphology
turns out basically to be useless and so can be scrapped doesn’t bother me at
all. Sticking to genes and voices, then, there are discrepancies among results
of different genetic markers used, proposed sister taxon pairs that are not
geographically closest relatives, and vocal analyses that focus on what I’m not
convinced aren’t trivial traits.
“I’m especially curmudgeonly about
vocal analyses, in general, but I’ll try not to go on too long and just sum up
my argument here as a general one, not directed specifically at this study.
It’s telling to me that DNA analyses use models of DNA evolution to weigh in on
what’s more or less important, what’s homologous or not, what kinds of changes
tend to be correlated, how to settle conflicting results, what genes to study,
etc. There’s no equivalent in vocal analyses. It’s just: measure a bunch of
stuff (usually the same stuff measured by somebody else citable) and then
decide on a statistical approach to what’s the same and what’s different and
how many differences or how much difference should be associated with distinct
species. I don’t think it should work that way or that vocal variation
associated with speciation evolves that way. There have to be considerations of
vocal context, repertoire, what kinds of traits vary and how, within individual
vs within population vs among populations comparisons, sexual variation, etc.
For example, is a faster or higher “tweet-toit-tweet” more different from a
slower or lower “tweet-toit-tweet” than it is from a “tweet-tweet-toit” song?
(In other words, do frequency or pace matter more than syntax?) Most vocal
analyses seem to me to suffer from the same kind of bean-counting attempts to
quantify qualitative or categorical differences that Brazilian job searches do
by using a point scale for everything on a person’s CV and then choosing the
candidate with the most points. Almost guaranteed to miss the best candidate
or, in the case of voices, not to recognize the real and relevant sources of
variation. So, in this case, without a clearer notion of how it was done and
what the sampling looks like, I can’t tell what’s diagnostic and how diagnostic
it really is.
“I guess that’s my biggest
conclusion, be it vocal or molecular or a combination, I’d like to see made
explicit what the diagnostic characters are for each taxon and to be convinced
that they really are diagnostic and that they’re meaningful traits.
“It’s especially disconcerting to
be a nay-sayer in the face of such a nice, big study. I suspect it’s a very good study! But I’m
unable to conclude anything very strongly from it. That may reflect my own
limitations. I’m perfectly willing to assume that the methods and sampling and
results are all correct, no mistakes, no interpretation problems. Even so, when
sister taxa are not neighbors or occupy different habitats or vocally
“identical” populations are separated geographically by others (as I believe was
found in this study), that sends up a red flag. Assuming those results are
correct, then there’s something very different going on from what’s found in
most groups—unusual and interesting—that seems to me to demand explanation. In
that case, identifying and understanding the processes that lead to such
atypical patterns seems more important than trying to force some taxonomic
conclusions on the situation.”
Comments
from Zimmer:
“Note that the reason I hadn’t voted on Part D of this Proposal previously, was
due to confusion over the way Part D was worded. I still haven’t seen any clarification as to
how to interpret the question being asked in Part D, which, based upon the
responses of others, seems to be an open question. Therefore, I am attempting to spell out my
vote in a way that it can be properly applied depending on what the question is
actually addressing.
“D. Treat intercedens as a separate
species from S. turdina. YES
to treating intercedens as separate from S. turdina, but NO to
treating it as a distinct species from S. amazonum. Given the evidence presented, I think it
needs to be recognized at the subspecific level (not as a synonym of amazonum),
but not as distinct at the species level from amazonum, wallacii
and cracrafti, all of which I would support as treating as part of a
polytypic S. amazonum.”