Proposal (1039) to South American Classification Committee
Split Camptostoma
obsoletum into six species
Background:
Camptostoma
obsoletum
currently comprises 13 subspecies taxa distributed across Central and South
America, from Costa Rica to Uruguay. Fragmentary vocal and genetic evidence
suggests that it may include multiple biological species (Ridgely and Tudor 1994, Ridgely and Greenfield 2001,
Rheindt et al. 2008). However, current taxonomy remains entirely based
on subtle variations in plumage and morphology, which are unlikely to reflect
true species limits in this group (Fitzpatrick 2004).
New
Information:
A
newly published analysis of geographic vocal variation in the genus Camptostoma,
based on 1,113 recordings of vocalizations from 994 individuals across 783
localities, identified six vocally distinct populations within C. obsoletum
(Lima and Vaz 2024). These
populations exhibit diagnostic differences in multiple different vocalization
types, including vocalizations that are typically species-specific in the
Tyrannidae. The populations are mostly allopatric but have abutting
ranges over large areas, with little evidence of gene
flow between them (Lima and Vaz 2024).
Assuming that the several diagnostic differences in multiple
vocalization types have the potential to generate substantial premating
isolation among these vocally distinct populations, and considering the
apparent pattern of parapatry without gene flow between several of these
populations, Lima and Vaz (2024) proposed splitting C. obsoletum into six
biological species, maintaining vocally undifferentiated taxa with potential
plumage differences as subspecies pending further study:
· Camptostoma
flaviventre Sclater & Salvin, 1865
o
C. f. flaviventre Sclater
& Salvin, 1865
o
C. f. majus Griscom, 1932
o
C. f. orphnum Wetmore, 1957
· Camptostoma
caucae Chapman, 1914
· Camptostoma
pusillum (Cabanis & Heine, 1860)
· Camptostoma
sclateri (Berlepsch & Taczanowski, 1884)
o
C. s. sclateri (Berlepsch
& Taczanowski, 1884)
o
C. s. maranonicum Carriker,
1933
o
C. s. griseum Carriker, 1933
· Camptostoma
napaeum (Ridgway, 1888)
o
C. n. napaeum (Ridgway, 1888)
o
C. n. olivaceum
(Berlepsch, 1889)
· Camptostoma
obsoletum (Temminck, 1824)
o
C. o. obsoletum (Temminck,
1824)
o
C. o. cinerascens (Wied,
1831)
o
C. o. bolivianum Zimmer,
1941
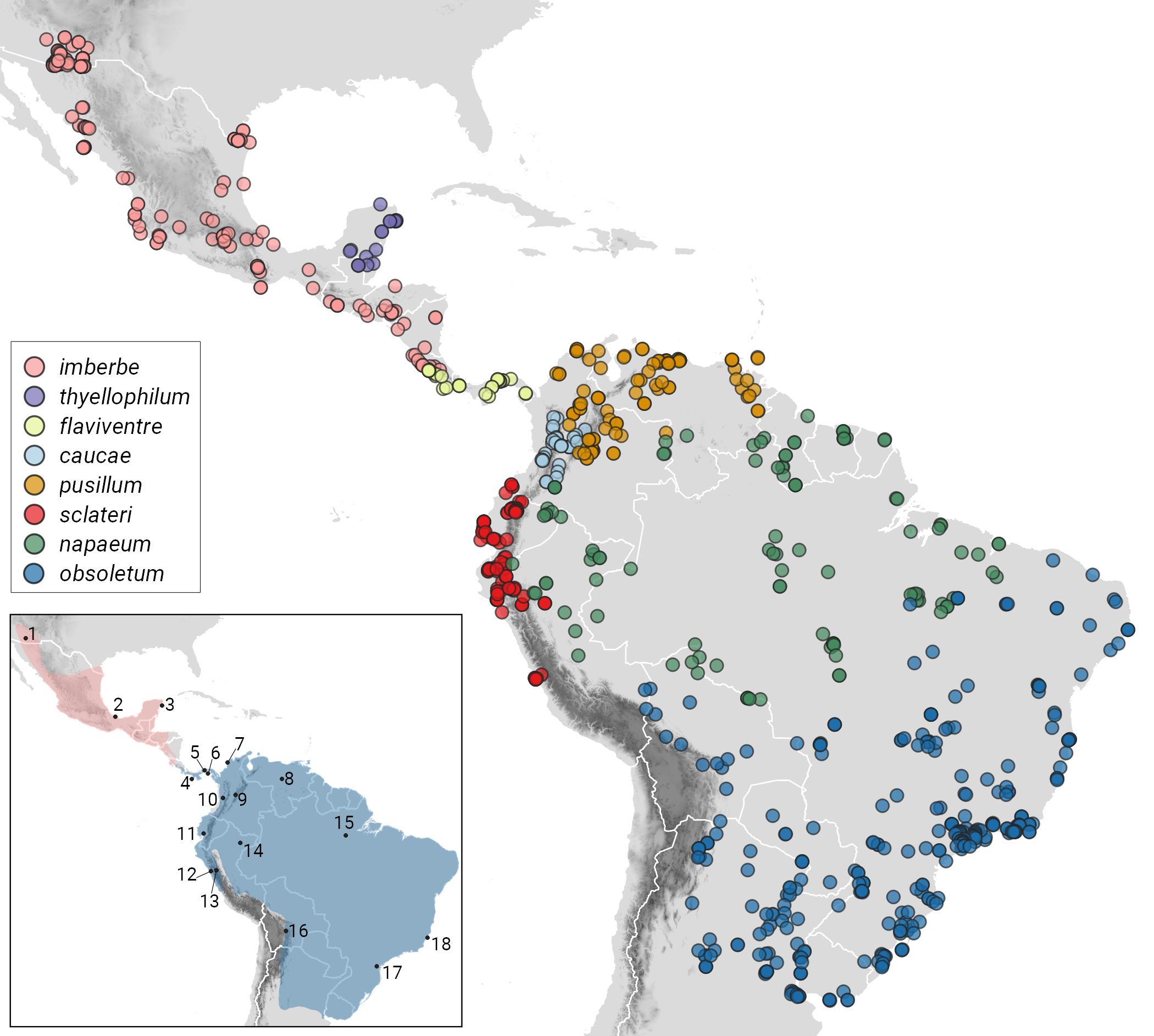
Here’s a figure from Lima and Vaz (2024) illustrating the
geographic ranges of the proposed species based on the sound recordings
examined. For further details about the figure, including information about
extralimital taxa, see Lima and Vaz (2024).
The proposal is subdivided as follows:
A. Treat sclateri (including maranonicum and griseum)
as a separate species from C. obsoletum
Camptostoma o. sclateri differs diagnostically from all
other taxa except maranonicum and griseum in daytime and dawn
songs as well as multiple calls (Lima and Vaz 2024). Although field playback
experiments suggest weak behavioral discrimination between sclateri and napaeum
based on daytime songs (Freeman et al. 2022), no evidence of hybridization
exists despite potential parapatry (Lima and Vaz 2024). Other vocalizations may
mediate premating isolation, although this remains untested (Lima and Vaz
2024). Alternatively, sclateri and napaeum may form undetected,
narrow hybrid zones.
B. Treat pusillum as a separate species from C.
obsoletum
Camptostoma o. pusillum differs diagnostically from all
other taxa multiple vocalizations, including daytime and dawn songs (Lima and
Vaz 2024). There is no evidence of gene flow with adjacent populations (caucae
and napaeum), despite potential parapatry (Lima and Vaz 2024).
C. Treat caucae as a separate species from C. obsoletum
Camptostoma o. caucae differs diagnostically from all
other taxa in daytime and dawn songs and multiple calls (Lima and Vaz 2024). No
evidence of hybridization exists with the apparently parapatric pusillum
and napaeum, suggesting strong premating isolation (Lima and Vaz 2024).
D. Treat napaeum (including olivaceum) as a separate
species from C. obsoletum
Camptostoma o. napaeum differs diagnostically from all
other taxa but olivaceum in multiple vocalizations, including daytime
and dawn songs (Lima and Vaz 2024). While differences with parapatric obsoletum
are subtle, and some hybridization may occur, phenotypic distinctiveness
despite extensive contact (~3,300 km) suggests substantial—albeit
incomplete—reproductive isolation (Lima and Vaz 2024).
E. Treat flaviventre (including majus and orphnum)
as a separate species from C. obsoletum
Camptostoma o. flaviventre, C. o. majus, and C. o.
orphnum collectively form a vocally unique population, differing
diagnostically from all others in multiple vocalizations (Lima and Vaz 2024).
Although this population occurs only in Central America based on the sound
recordings examined by Lima and Vaz (2024), I included it here in the proposal
because it may plausibly occur in extreme western Colombia near Panama. I defer
to the committee to determine whether South American records exist.
References:
Fitzpatrick,
J. W. (2004). Family Tyrannidae (Tyrant-flycatchers). In Handbook of the
Birds of the World (J. del Hoyo, A. Elliott and D. Christie, Editors). Lynx
Edicions, pp. 170–462.
Freeman, B.
G., J. Rolland, G. A. Montgomery, and D. Schluter (2022). Faster evolution of a
premating reproductive barrier is not associated with faster speciation rates
in New World passerine birds. Proceedings of the Royal Society B
289:20211514.
Lima, R.
D., and R. V. Vaz (2024). Divergence in vocalizations indicates cryptic
speciation in Camptostoma tyrannulets. Ornithology: ukae058[RL1] .
Rheindt, F.
E., J. A. Norman, and L. Christidis (2008). Genetic differentiation across the
Andes in two pan-Neotropical tyrant-flycatcher species. Emu 108:261–268.
Ridgely, R.
S., and P. J. Greenfield (2001). The birds of Ecuador: status, distribution,
and taxonomy. Cornell University Press.
Ridgely, R.
S., and G. Tudor (1994). The birds of South America. Volume 2 (The Suboscine
Passerines). University of Texas Press.
Rafael D. Lima, December
2024
Note
from Remsen on English names: A separate proposal on English names will be
needed if any of these proposals passes.
___________________________________________________________________________________________________
Vote tracking chart: https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Lane:
“YES. Regardless of whether there is a molecular component to this study, I
applaud the authors’ efforts to break down a potential headache of a complex,
and I see this study as good enough to act upon here. I agree with their
assessments of the taxa, the vocal groupings, and their outlining of daughter
species. YES to all proposed splits from C. obsoletum.”
Comments
from Zimmer:
“YES to all splits (see separate subproposals below) as advocated in the
Proposal, and as suggested by Lima & Vaz 2024. I have long grappled to make sense of what I
was perceiving as vocal distinctions within the C. obsoletum group
across its broad range, and have it mesh with plumage differences and
understood distributions of named taxa, but without the kind of geographically
broad-based and well-sampled vocal analysis conducted by Lima & Vaz (2024),
it has remained a muddled mess in my mind.
Part of the confusion is due to the effects of wear and fading on some
of the few plumage characters that can actually allow visual discrimination of
taxa in the field, and part of it is due to each population having multiple
types of vocalizations in their repertoires (songs, dawn songs, contact calls,
agitation/interaction calls, etc), and without a systematic, thorough analysis,
you can’t be sure that you are always comparing apples to apples and oranges to
oranges. The absence of a molecular
component to this study does not bother me, since I believe vocal differences
in these phenotypically conserved groups of suboscines are almost always going
to provide the strongest indication of species-limits.
“A. Treat sclateri (including maranonicum
and griseum) as a separate species from C. obsoletum: YES, also matches my
field impressions of vocal distinctions.
“B. Treat pusillum as a
separate species from C. obsoletum: YES, also matches my field
impressions of vocal distinctions.
“C. Treat caucae as a separate species
from C. obsoletum: YES, based upon vocal distinctions and no evidence of
hybridization with parapatric pusillum and napaeum, as detailed
in Lima and Vaz 2024.
“D. Treat napaeum (including olivaceum)
as a separate species from C. obsoletum. YES. This also matches my field impressions
of vocal distinctions.
“E. Treat flaviventre (including majus
and orphnum) as a separate species from C. obsoletum. YES. This one with
particular enthusiasm, based on my own field impressions in Panama/s Costa
Rica. I do not, however, have any
evidence for whether South American records of flaviventre exist.”
Comments from Areta: “I´ve been amassing recordings of Camptostoma for many years now, which
match most of Lima & Vaz´s conclusions. The only point of discontent is at
the junction of flaviventre, caucae, and pusillum, which seem vocally very similar in some respects but
apparently not in others. I am uncertain on whether the daysong of caucae is really the one chosen by the
authors, instead of the slower, more whistled vocalization; likewise, I am
unable to hear ANY consistent distinction between dawnsongs of flaviventre (https://macaulaylibrary.org/asset/60355 https://macaulaylibrary.org/asset/57359 https://macaulaylibrary.org/asset/147441041) and caucae (https://xeno-canto.org/307322 https://xeno-canto.org/344295 https://macaulaylibrary.org/asset/100724081). The sample sizes are
small, which can be tricky in the highly variable Camptostoma dawn songs. I´ve also found some other issues around flaviventre, pusillum, and caucae: see
for example this dawn song of pusillum
from Zulia (Venezuela) https://macaulaylibrary.org/asset/69881, and this dawn song
attributed to caucae from
Antioquia (Colombia) https://macaulaylibrary.org/asset/100724081, both of which share a
distinctive inflection in the second and third note and sound remarkably alike,
and also add this flaviventre in the
mix https://macaulaylibrary.org/asset/25603. This one from the
Coastal Cordillera of Venezuela seems also quite similar to songs of flaviventre: https://macaulaylibrary.org/asset/69878. Also, these trills
from pusillum (https://macaulaylibrary.org/asset/557328701 https://macaulaylibrary.org/asset/530472281 https://macaulaylibrary.org/asset/34477 https://macaulaylibrary.org/asset/478141441) and these of caucae (https://macaulaylibrary.org/asset/283449281 https://macaulaylibrary.org/asset/272722131 https://macaulaylibrary.org/asset/629232892). These voices of pusillum https://macaulaylibrary.org/asset/597111551 are much like this one
from a zone of presumed caucae https://macaulaylibrary.org/asset/534542081 which in turn is very
similar to this one of caucae https://macaulaylibrary.org/asset/626683461, and this one of flaviventre https://macaulaylibrary.org/asset/67231361.
“Given
the broad similarities, I am not convinced that flaviventre, caucae and pusillum are different species, and so I
vote to recognize them under a single species: C. pusillum, or
alternatively, split pusillum and
leave flaviventre and caucae as one thing (which also seems
problematic). The most striking difference in vocalizations seems to be the
trill that has been attributed to caucae,
and which seems to be missing from other populations. Is this vocalization
really from Camptostoma? Has this
gone unrecorded in flaviventre? After
hearing a fair share of recordings unsystematically, I came away unconvinced of
the vocal distinctions, especially those between caucae and flaviventre.
Maybe misidentifications to taxon are playing badly on us here, but I can
easily imagine the vocal types transitioning through geography (e.g., based on
what I hear, some pusillum sound
slightly higher-pitched and somewhat faster than caucae and flaviventre,
especially if one compares northern to southern examples), and the similarities
seem to me to carry more weight than the differences. I don´t think that the
spectrogram of the diurnal song of flaviventre
is really representative of the taxon: in listening to all the daytime songs
assigned to this category by Lima & Vaz, I hear a lot of variation, most of
which do not fit the spectrogram. I am not fully convinced of anything of the
above, but I also do not see strong evidence to split the three as different
species. I would like to hear what others think on the northern South America-southern
Central America issue.
“Meanwhile,
my votes are:
A. Treat sclateri (including maranonicum and griseum)
as a separate species from C. obsoletum --- YES
B. Treat pusillum as a separate species from C. obsoletum --- YES
C. Treat caucae as a separate species from C. obsoletum --- YES (separate from C.
obsoletum) and NO (to treat is as a species on its own): treat caucae
as subspecies together with C. pusillum or together with flaviventre.
D. Treat napaeum (including olivaceum) as a separate
species from C. obsoletum --- YES
E. Treat flaviventre (including majus and orphnum)
as a separate species from C. obsoletum --- YES (separate from C. obsoletum) and NO (to
treat is as a species on its own): treat flaviventre as subspecies
together with C. pusillum or together with caucae.”
Comments
from Niels Krabbe (voting for Del-Rio): “After reading Nacho's comments, I am in
doubt whether pusillum, caucae, and flaviventre are
separate species, but YES to the rest of the suggested splits. So:
“A: YES. Treat sclateri
(including maranonicum and griseum) as a separate species from C.
obsoletum
B: YES. Treat pusillum
as a separate species from C. obsoletum [but including the subspecies flaviventre,
caucae, majus, and orphnum]
C: YES and NO. Treat caucae
as a separate species from C. obsoletum [but as a subspecies of pusillum]
D: YES. Treat napaeum
(including olivaceum) as a separate species from C. obsoletum
E: YES and NO. Treat flaviventre
(including majus and orphnum) as a separate species from C.
obsoletum [but as a subspecies of pusillum]”
Additional
comments from Rafael Lima: “I do not have field experience with the northern taxa,
so I cannot confirm with certainty that the rapid trill belongs to Camptostoma.
Our assumption was based on two points. First, multiple experienced observers
and ornithologists have recorded this vocalization and consistently attributed
it to Camptostoma. Second, if the slower, more whistled vocalization
in Figure 9H represents the daytime song of caucae, then there is no
clear homologous counterpart among similarly structured vocalizations in flaviventre
and pusillum (Fig. 9G–I). However, the key point remains:
as noted in the paper, regardless of whether the daytime song of caucae is
the rapid trill (Fig. 2D) or the slower, more whistled vocalization (Fig. 9H), caucae
still has a distinctive daytime song. In other words, the conclusion that caucae
has a unique daytime song remains unchanged, irrespective of this analytical
choice. (All data and scripts necessary to replicate the analyses and figures
are publicly available at https://datadryad.org/dataset/doi:10.5061/dryad.612jm64dm, allowing for
alternative analyses to test whether different methodological choices affect
the conclusions.)
“Regarding
the dawn songs of flaviventre, pusillum, and caucae, they
are indeed similar. While small sample sizes for flaviventre and caucae
introduce some uncertainty, the subtle differences in their dawn songs were
consistently captured in the measurements of the main phrase (Fig. 5),
suggesting they are not spurious. However, even if one assumes that these dawn
song differences are artifacts of sample size limitations, the distinctiveness
of their daytime songs remains.
“Beyond
the magnitude of vocal differences, the apparent pattern of widespread
parapatry without gene flow between caucae and pusillum further
supports their recognition as separate species (as discussed in the paper). The
case for treating pusillum as distinct from flaviventre is less
strongly supported, as it relies on the (subtle) vocal differences alone.
Still, I consider the differences in their daytime songs alone sufficient to
justify treating all three taxa as separate species.”
Comments
from Tom Schulenberg (voting for Jaramillo): “YES -- accept all of the splits, as
proposed by Lima and Vaz. I should say that the taxa with which I have the
least experience - flaviventre, caucae, and pusillum - are
at the heart of the issues here; splitting all other taxa is straightforward. I
acknowledge Nacho's concerns, and I'll admit that the differences between flaviventre,
caucae, and pusillum in some vocalizations are subtle. I too am
surprised at the trill attributed to caucae, but I'm willing to accept
that this vocalization is from a Camptostoma. it seems to be a frequent
enough vocalization, and if it's not Camptostoma, then ... what is it? I
wouldn't set it aside without having a clear alternative contender, which in
this case apparently would need to be a species whose distribution corresponds
to that of caucae. So, taking this trill as caucae, and in the
absence of evidence for this vocalization in other populations, then I don't
see any alternative other than to split this taxon.”
Comments
from Stiles:
“YES to all splits as proposed after reading Lima's comments regarding
vocalizations.”
Comments
from Remsen:
“YES to all parts. The paper and
proposal do a great job summarizing and synthesizing existing data on vocal
differences. Although this might not be the
final word on species limits in the group (as per Nacho’s comments), it is
certainly a major step forward and a great improvement over the status
quo. I can’t really add anything
specific to all the comments above.”
Comments
from Robbins:
“I spent a fair amount of time listening to vocalizations of this complex over
the course of two different occasions. Based on the vocalizations, three of the
splits are indeed straightforward. Now,
based on Rafael’s response to good points made by Nacho, I feel comfortable
with recognizing all proposed splits, i.e., YES to A-E. Kudos to Lima and Vaz for sorting this out.
Naturally, we all look forward to a genetic assessment of the complex.”
Comments
from Claramunt:
“NO. Levels of genetic divergence and the diversity of
vocalization suggest that more than one species may be involved. The study by Lima and Vaz has an
impressive sample size and good coverage of all subspecies. However, an
important piece of evidence is missing: whether the
geographic variation is discrete and not clinal. In some instances, the study
argues that no intermediate birds were found in putative contact zones, but a
formal analysis assessing whether variation is discrete (a step-cline) or
gradual-clinal is missing. It’s easy to find “statistically significant
differences” between the means of predetermined groups, but that is not
sufficient evidence because a gradual cline can show statistically significant
differences between arbitrary subdivisions of it. Trends should be analyzed
instead with standard spatial regression models, cline analysis methods, or
Normal Mixture Models (Cadena et al. https://doi.org/10.1093/sysbio/syx071).
Response
from Rafael Lima:
“Santiago's concern, that the species-level taxa proposed in the paper could be
artifacts of dividing gradual clines, is a valid one. Although we did not
explicitly show this in the paper, I want to clarify that there is no
noticeable geographic vocal variation (clinal or otherwise) within any of the
eight species-level taxa. Importantly, we did not rely on predetermined groups
like subspecies labels from recordings. As described in the methods,
populations were delineated based on visual differences in spectrograms, so the
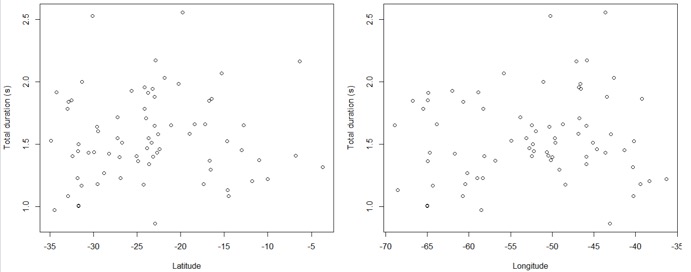
patterns emerged organically from the data. To illustrate that there is no
clinality, consider the total duration of daytime songs of obsoletum.
This trait is variable (see scatter in Fig. 3B) across a widely distributed taxon, but plotting it against latitude or longitude shows
no association with geography. Similar plots can be generated for all acoustic
variables and taxa using the data deposited in Dryad. One can also visualize
variables on maps to see the lack of consistent variation within the eight
taxa. Put simply, the variation within each taxon is individual rather than
geographic and, since there is not even geographic variation within these taxa,
there is no clinality among them.”
Response
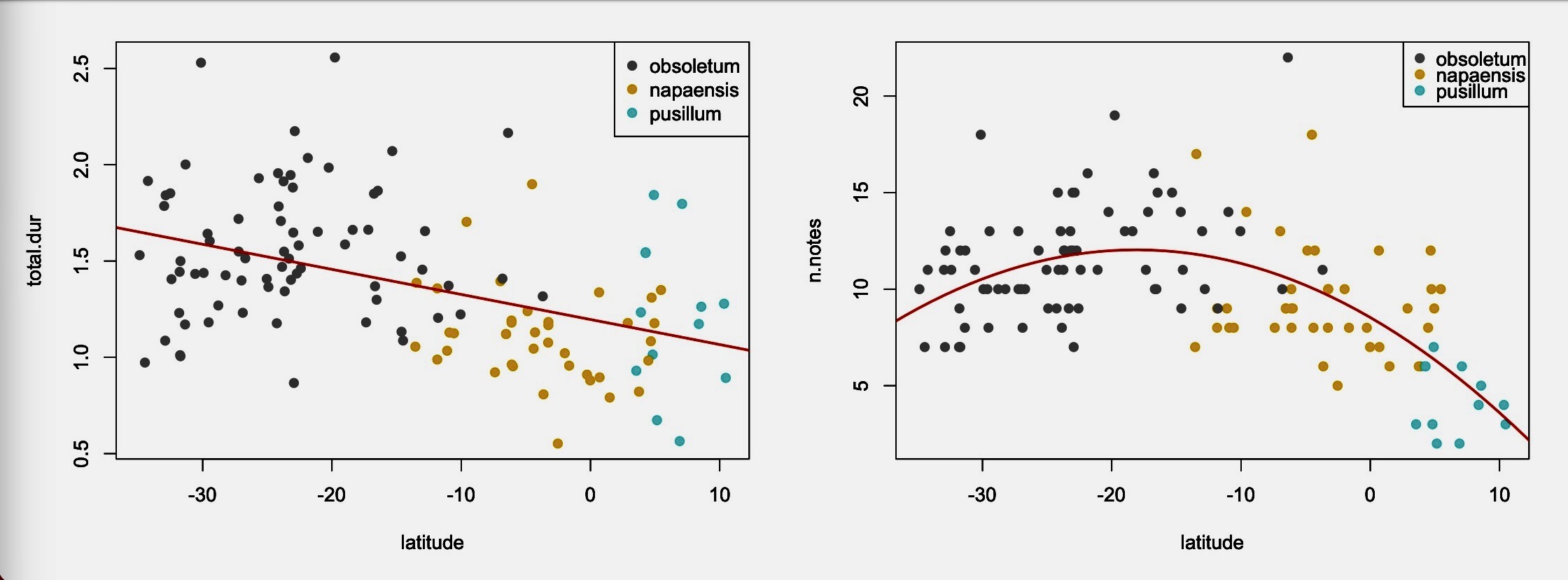
to Lima from Claramunt: “The relevant clinal
variation I’m referring to is not within taxa but across taxa. See for example
the attached plot of Total daysong duration for the
three taxa in the South American lowlands. Within obsoletum, a cline is
not detectable, but variation across the three taxa looks like a gradual cline
in which total duration decreases gradually towards the north. No
discontinuities or step as apparent, suggesting clinal variation that may be
the product of gene flow, a climate or latitudinal selective pressure, or both.
For the number of notes there seem to be a nonlinear trend across the
continent. The fact that pusillum has a ‘significant’ reduction in the
number of notes is immaterial, as it could be explained by being at one extreme
of a continental-wide gradual trend.