Proposal (1055) to South American Classification Committee
Revise species limits
in Onychorhynchus coronatus.
Effect
on SACC:
This could split widespread Onychorhynchus coronatus into as many as six
species.
Background: Our current Note is
as follows:
2. Ridgway (1907), Cory & Hellmayr
(1927), and Pinto (1944) considered the four
subspecies groups in Onychorhynchus coronatus as separate
species: mexicanus of Middle America and northwestern Colombia, occidentalis
of western Ecuador and northwestern Peru, coronatus of Amazonia, and swainsoni
of southeastern Brazil. Meyer de Schauensee (1966, 1970) treated them all as
conspecific without providing justification, and this was followed by Traylor
(1977<?>, 1979b), AOU (1998), Sibley & Monroe (1990), Fitzpatrick
(2004), Ridgely & Tudor (1994), who provided rationale for their continued
treatment as conspecific, and Dickinson & Christidis (2014), but this was not
followed by Wetmore (1972), who considered the evidence insufficient for the
broad treatment. Ridgely & Greenfield (2001) and Hilty (2003) returned to
the classification of Cory & Hellmayr (1927). Collar et al. (1992)
considered occidentalis as a separate species. See Whittingham &
Williams (2000) for analysis and discussion of morphological characters. Del
Hoyo & Collar (2016) recognized four species: O. mexicanus of Middle
America and n. South America; O. occidentalis of the Tumbesian region; O.
coronatus of Amazonia; and O. swainsoni of the Atlantic Forest. Reyes et al. (2023) presented data relevant
to recognition of as many as six separate species based mainly on deep
divergence in mtDNA. SACC proposal badly needed.
Birds
of the World/Clements has instituted a 2-way split: https://ebird.org/species/royfly1
Ridgely
& Tudor (1994) thought that perhaps four species could be recognized, but
that treating them all as conspecific was the best course given similarities I
behavior and voice, as far as was known at the time. (They mistakenly cited AOU 1983 as having
recognized two species.)
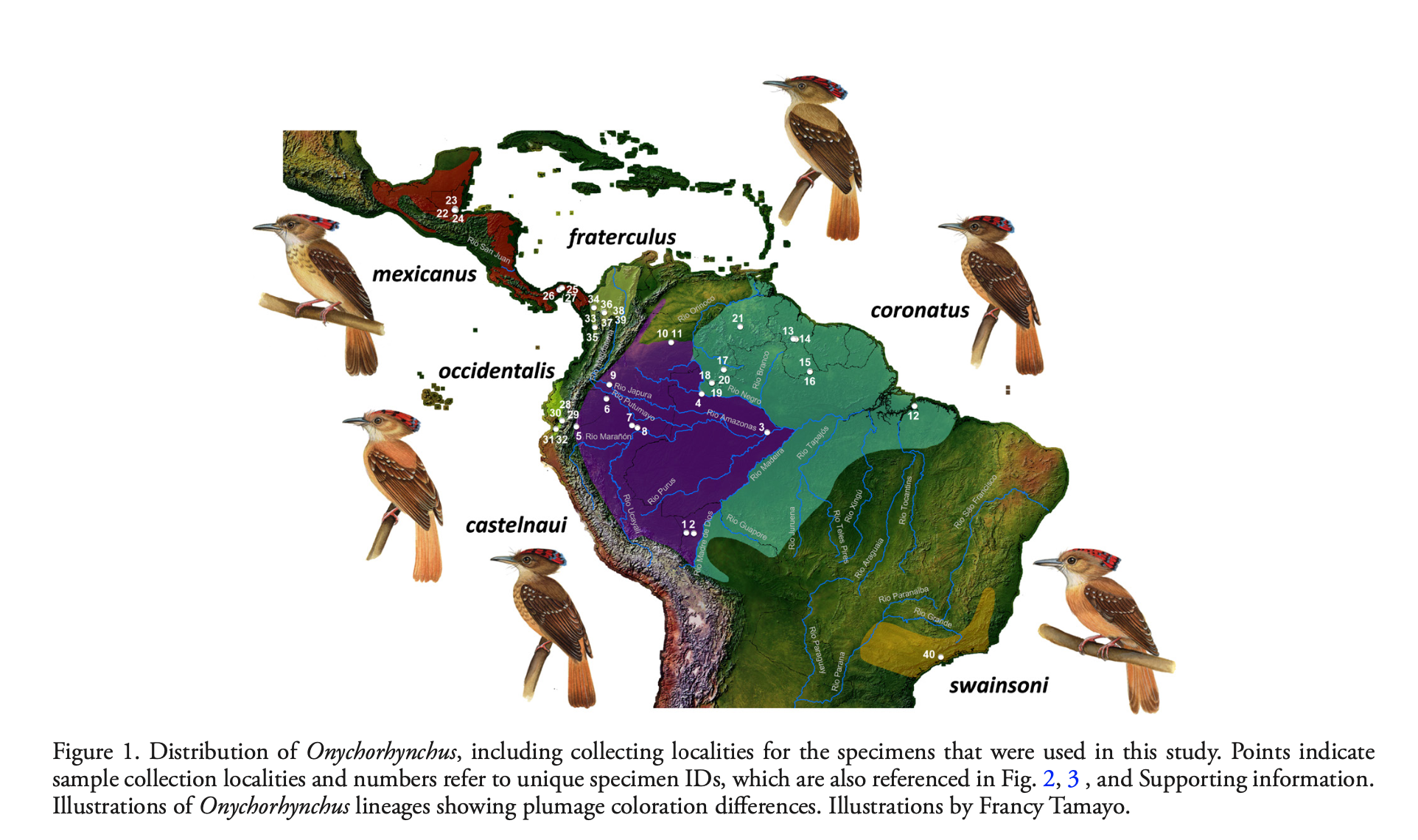
Six
taxa are recognized in the complex (Dickinson & Christidis 2014):
• mexicanus (s. Mexico to Panama)
• fraterculus (n. Colombia and nw.
Venezuela
• occidentalis (Tumbesian region)
• coronatus (Guianan Shield and n. and
e. Amazonian Brazil)
• castelnaui (w. Amazonia)
• swainsoni (se. Brazil)
Here
are Hilary Burn’s illustrations from del Hoyo and Collar (2016), which
illustrate the differences in plumage, primarily in degree of markings on
breast and general plumage tone. Note
that swainsoni and geographically distant occidentalis are the palest
and least spotted of the group, and I suspect this is what influenced Meyer de
Schauensee’s reasoning for treating them all as conspecific.

In
Harvey et al.’s (2020) massive phylogenetic analysis of the suboscines using
genomic (UCE) data, 4 subspecies were included.
Swainsoni was “basal” to the other taxa, with an estimated
divergence time of ca. 6 MYA; nominate coronatus was the sister to mexicanus
+ occidentalis, with divergence estimate of ca. 3 and ca. 1 MYA,
respectively:

Just
eye-balling node depth in adjacent clades in the Harvey et al. tree indicates
that 6 MYA divergence is more typical for taxa treated at the species level,
whereas 3 and 1 MYA are more typical for taxa traditionally treated as conspecific. This is no substitute for a formal analysis
but is meant only to provide a crude comparison of node depths. Feel free to make your own comparisons, of
course.
New
information:
Reyes et al. (2023; note that the “et al.” includes our Luciano and Elisa)
produced the first genetic analysis of the complex. They used a single mtDNA marker: NADH. They sampled 40 individuals, including
individuals of all taxa in the complex.
They had only 1 swainsoni but had at least 5 for the other 5
taxa.
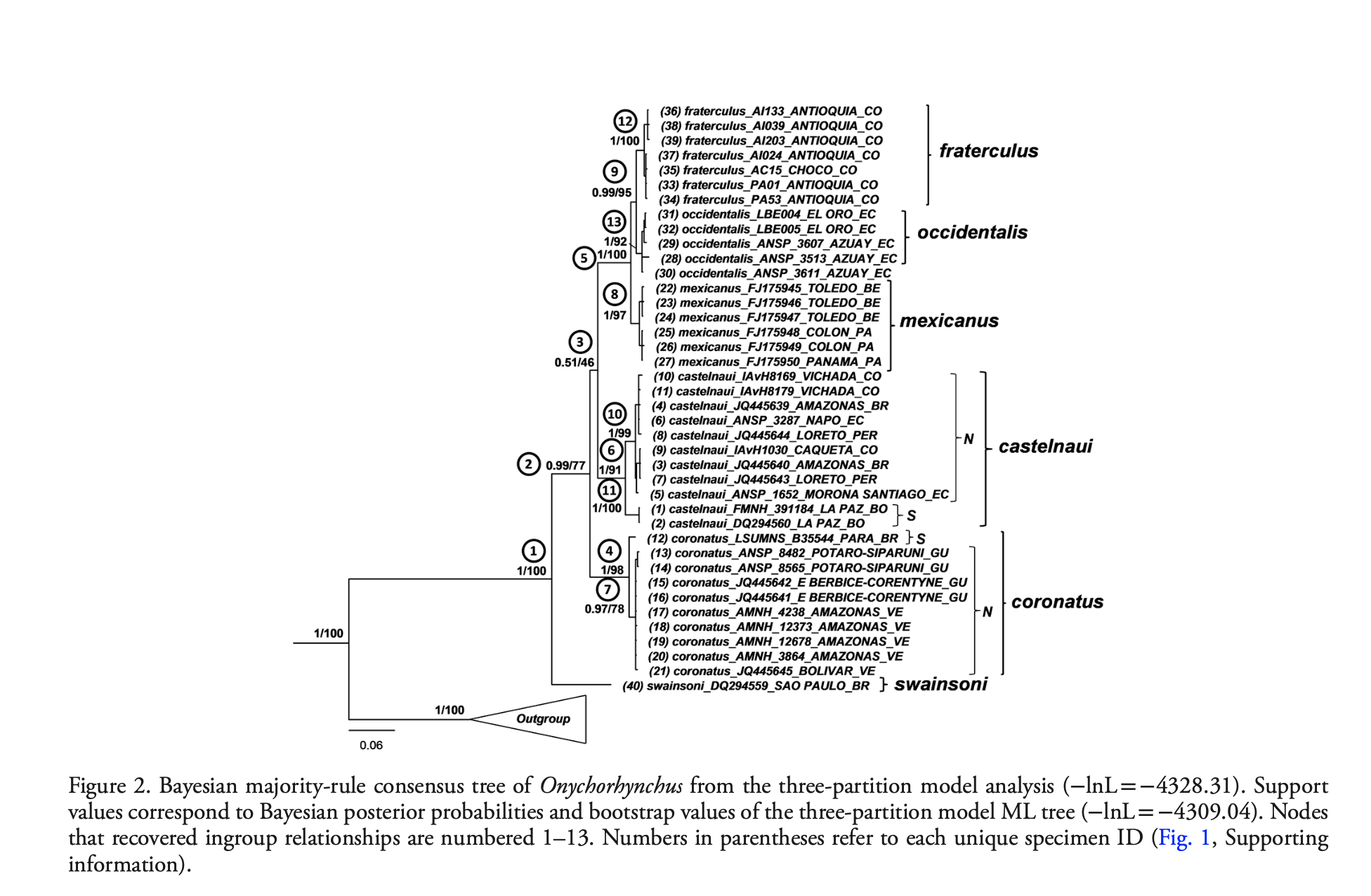
The
topology of the tree was consistent with that of Harvey et al. (2020),
including the placement of the taxon with arguably the most divergent plumage, occidentalis,
within the northern mexicanus group.
Their geographic sampling was reasonably thorough except for the absence
of specimens from the populations attributed to coronatus from Brazil
south of the Amazon and e. Bolivia:
The
mean estimate of the divergence time between swainsoni and the rest was
6.1 MYA, remarkably similar to that of Harvey et al. (2020). The focus of the paper was on historical
biogeography, not taxonomy, so the paper did not produce any firm taxonomic
recommendations. Nevertheless, their
analysis produced evidence for 6 independently evolving mtDNA lineages that
they suggested could be treated as species.
Caution is required because the geographic and numerical sampling is possibly
insufficient to be certain that the lineages are monophyletic, at least in
southwestern Amazonia, where large areas of coronatus distribution in
potential contact with castelnaui have not been sampled. Their
concluding paragraph is as follows:
“Furthermore,
this study helped to reveal independently evolving lineages that might have to
be treated as separate species with different conservation concerns.
Complementary studies that include nuclear DNA, morphology, niche
differentiation, and vocalizations with thorough sampling throughout the Onychorhynchus
distribution are needed to fully resolve the evolutionary relationships and
delimit species within this genus.”
Potential
changes in species limits are also implied in the section titled Cryptic
Diversity. I have to point out that the
diversity cannot be described as “cryptic” if it has been known for more than a
century with at least 4 taxa treated as separate species for the first half of
the 20th century, and the only taxon not described before 1860 was fraterculus
Bangs 1902. In fact, Reyes et al. make
this very point in that same section.
As
for vocalizations in this group, analyses are handicapped by how few samples
there are. Royal Flycatchers vocalize
infrequently, and dawn songs are poorly known.
Thus, It is no surprise to me that Peter Boesman did not include this
species as one of his 400+ “Ornithological Notes” (for HBW/BLI) despite this
complex being a prime candidate for a preliminary analysis.
Kirwan
et al. (2024a, b) summarized what is known about vocalizations in this group,
with links to recordings. In Birds of
the World, they are treated as two species: O. swainsoni for se. Brazil
and O. coronatus for everything else.
Kirwan et al. (2024b) outlined the differences used by del Hoyo &
Collar’s (2016) adoption of the Tobias et al. scoring system and said this
about voice: “Also distinctive in call, which is significantly shorter (effect size
4.9; score 2) and higher-pitched (effect size also 4.9; score 2) than that of
other taxa.” [It sounds lower-pitched to me.] Here are the
recordings cited by Kirwan et al. (2024b):
https://search.macaulaylibrary.org/catalog?taxonCode=royfly5&mediaType=audio&sort=rating_rank_desc
Kirwan
et al. (2024a) described the vocalizations of the coronatus group as
follows:
“Song. A
rarely heard series of whistles, which apparently varies geographically. There
are however hardly any recordings available, thus the following are merely
examples of specific cases:
· Northern
group: audio 2 In
Costa Rica and Panama, a series of rather sharp downslurred whistles preceded
by a short introductory note, whit..eeeuw...eeeuw...eeeuw ..., uttered
at a rate of about one whistle per second. In the literature, Song in this
region is described as a long series of higher, sharper notes with a most
peculiar intonation (8) or
(in Mexico) a descending, slowing series of plaintive whistles, usually 5‒8, whi'
peeu peeu peeu peeu peeu ..., or wh' wheeu wheeu ... (65).
· Amazonian
group: In Brazil, a series of long melodious whistles, starting with a loud
flat-pitched introductory note and followed by a series of lower-pitched
disyllabic mellow whistles wheeee-pihuuw-pihuuuw-pihuuw.
Another variant is structurally similar, but disyllabic whistles are more
modulated wheeeee-priririuuw-priririuuw... audio Also
described as a squeaky PEE'u occasionally followed by a lower,
musical PEE'u-brrrr (66).
· Pacific
group: Possibly a homologous vocalizations is
described as a squeaky whi-CHEW in a series in a display (66). No
recordings of Song are available.
“Primary call. audio 3 A
short disyllabic nasal keeeyup, repeated many times at intervals of
ca. 2 seconds. On sonogram, note has a characteristic shape, initially reaching
a flat-pitched top around 3 kHz, after which frequency drops sharply with a
hiccup around 2 kHz, for a total duration of about 0.20‒0.25 second. Also
described as a low-pitched sur-líp, sometimes [repeated] over and
over (67); a
loud, mellow, hollow-sounding whistle, usually two-syllabled: keeeyup or
keee-yew (8); a
squeaky to hollow, plaintive whee-uk or see-yuk (65);
and as a loud, plaintive squeak: PEE'yuk (42).”
Three
recordings of the primary call are presented here, one from mexicanus,
one from castelnaui, one from coronatus: https://birdsoftheworld.org/bow/species/royfly1/cur/sounds#vocal. These indeed sound very different from swainsoni,
and sound very similar to each other. No
primary call of occidentalis was presented.
Discussion
This
complex has been on everyone’s radar “forever” in terms of species limits,
especially because classifications in the early 1900s treated them as four
species, and Meyer de Schauensee provided no rationale for the lump. One could argue on that basis alone that a
return to the taxonomy of Ridgway-Cory-Hellmayr-Pinto is warranted, and thus place
burden-of-proof on treatment as a single species.
Based
on qualitative perusal of sonograms of what is called the primary call, one
could easily justify treating swainsoni as a separate species, as done
by Birds of the World. Based on that
same qualitative perusal of N=1 primary calls of mexicanus, coronatus,
and castelnaui, the primary calls sound similar to me, and their
sonograms have that distinctive appearance noted by Kirwan et al. Calls of occidentalis and fraterculus
were not presented, but perusal of xeno-canto suggests that occidentalis
(N=1; https://xeno-canto.org/264700) and fraterculus (N=1; https://xeno-canto.org/664102)
are also qualitatively similar. However,
none of this is a substitute for a quantitative analysis, so that concerns me.
It
is not the job of SACC members to do original quantitative analyses, and this
is what would be ideal for a decision on species limits in Onychorhynchus. The qualitative difference between swainsoni
and the others could be considered sufficient, in my opinion, for placing
burden-of-proof on a single species treatment.
This is also consistent with the genetic data of Harvey et al. (2020)
and Reyes et al. (2023). However, in my opinion, further splits would require
quantitative analyses of the vocalizations.
Studies of contact zones would be the best of all, but occidentalis
is isolated, and I’m not sure if contact zones exist between mexicanus
and fraterculus, fraterculus and coronatus, fraterculus
and castelnaui, or coronatus and castelnaui.
For
voting purposes let’s break this down as follows, with YES/NO votes on the
following
A. Retain traditionally
defined broad O. coronatus. If
YES, then B, C, and D are automatically NO.
B. Recognize two
species (as in HBW and Birds of the World): O. swainsoni and O.
coronatus, which would include all other taxa.
C. Recognize four
species: O. mexicanus, O. occidentalis, O. swainsoni, and O.
coronatus (return to the classification of Cory & Hellmayr and others,
following del Hoyo and Collar 2016):
D. Recognize six
species (O. mexicanus, O. occidentalis, O. fraterculus, O. castelnaui, O. coronatus, and O. swainsoni) for each of the lineages
elucidated by Reyes et al. (2023).
Pending
input from those more knowledgeable than I, my recommendations are as follows:
A. No – the original lump was never justified; B- Yes – based on published
sonograms of primary call note and a genetic distance more consistent with
species rank in related lineages; C. No – this would require additional,
qualitative research on vocalizations in my opinion; D. No – likewise, and I
oppose using mtDNA lineages for taxonomy, particularly because of the
well-known potential problem of gene trees conflicting with species trees; this
point was implicit in Reyes et al.
Literature
Cited:
KIRWAN, G. M., R. SAMPLE, B. SHACKELFORD, R. KANNAN,
AND P. F. D. BOESMAN. 2024a. Atlantic Royal Flycatcher (Onychorhynchus
swainsoni), version 1.1. In Birds of the World (T. S. Schulenberg and B. K.
Keeney, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.royfly5.01.1
KIRWAN, G. M., R. SAMPLE, B. SHACKELFORD, R. KANNAN,
AND P. F. D. BOESMAN. 2024b. Atlantic Royal Flycatcher (Onychorhynchus
swainsoni), version 1.1. In Birds of the World (T. S. Schulenberg and B. K.
Keeney, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.royfly5.01.1
REYES, P., J. M. BATES, L. N. NAKA, M. J. MILLER, I.
CABALLERO, C. GONZALEZ-QUEVEDO, J. L. PARRA, H. F. RIVERA-GUTIERREZ, E.
BONACCORSO, AND J. G. TELLO. 2023. Phylogenetic relationships and biogeography
of the ancient genus Onychorhynchus (Aves: Onychorhynchidae) suggest
cryptic Amazonian diversity. J. Avian Biology 2023: e03159.
WHITTINGHAM, M. J., AND R. S. R. WILLIAMS. 2000. Notes on morphological differences
exhibited by Royal Flycatcher Onychorhynchus coronatus taxa. Cotinga 13: 14-16.
Note
on English names:
BOW uses “Atlantic Royal Flycatcher” for swainsoni and “Tropical Royal
Flycatcher” for everything else. If
Option B passes, then I think we can save ourselves plenty of work and simply
adopt the current BOW names Tropical Royal-Flycatcher and Atlantic
Royal-Flycatcher. If you object and
would like a separate proposal, and are also willing to write that proposal,
speak out. “Tropical” may only be a temporary
name pending more data on future splits.
There will be the usual howls concerning use of names of oceans as names
of terrestrial species, but I think it is widely understood what the
implication of such names are for land birds.
If Option C or D passes, then we will have to do a separate SACC
proposal on English names depending on which taxonomy we adopt. See the illustration above for names used by
BLI for a 4-way split.
Van Remsen, May 2025
Voting Chart: https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Robbins:
“Van has done an excellent job of distilling the issues with species limits
within coronatus. I agree with
his concluding assessment that more complete genetic sampling (it is
unfortunate that the Harvey et al. UCE data lacked two of the taxa) is needed,
especially from western Amazonia. For
reasons that fieldworkers have known for a long time and was reiterated by Van,
relevant vocal data with sufficient sample sizes may be a long time in
coming. Although there are certainly
more than two species involved, to be conservative at
this point, I follow Van’s recommendations:
“A. NO
B. YES
C. NO
D. NO”
Comments
by Lane:
“YES to B and NO to the other options until a better case is built for
additional splits. I have uploaded my recordings of occidentalis with
the “whi-chew” vocalization (ML637382910... cut off,
unfortunately), suggesting it to be similar to other members of the overall “tropical”
group. I can’t argue that the evidence to split this group up more is
convincing at this time, and thus vote for a 2-species taxonomy largely based
on vocal differences.”
Comments from Areta: “First of all, Reyes et al. (2023) indicated the need of further
sampling and the integration of more lines of evidence in order to sort the
taxonomy of O. coronatus. Thus, any
taxonomic decision taken at this point will not be the most informed one.
“Secondly, I see the phylogenetic tree
of Harvey et al. (2020) as being consistent with that in Reyes et al. (2023),
except that it lacks samples from some key taxa.
“Third, my vote for AviList/WGAC on this
issue before the publication of Reyes et al. (2023) was ‘Initially, assuming the topology and divergences shown in Harvey et al
2020 are representative of this complex, I was leaning towards a 3-way split
(Atlantic Forest, "Amazon", and Central America/Pacific South
America). But after listening to the relatively good sampling of calls across
the range, I agree with the 2-way split:
Atlantic Forest vs rest of the World. I hope someone undertakes a deeper
analysis on Onychorhynchus!"
A. Retain traditionally defined broad O. coronatus. NO
B. Recognize two species (as in HBW and Birds of the World): O.
swainsoni and O. coronatus, which would include all other taxa.
Maybe (see below)
C. Recognize four species: O. mexicanus, O. occidentalis, O.
swainsoni, and O. coronatus (return to the classification of Cory
& Hellmayr and others, following del Hoyo and Collar 2016). NO, this is
untenable in light of the phylogenetic results: how can we split occidentalis (with fraterculus) from mexicanus
without also splitting castelnaui
from coronatus?
D. Recognize six species (O. mexicanus, O. occidentalis, O. fraterculus, O. castelnaui, O. coronatus, and O. swainsoni) for each of the lineages elucidated
by Reyes et al. (2023). NO, this seems to be over-splitting the mexicanus group, which has shallow
divergences. The biogeographic pattern linking birds from NW South America and
the Pacific of Ecuador and Peru to birds from Central America-Mexico is not
unusual. Sometimes the taxa have speciated, sometimes they have not. In this
case, I don´t think the level of divergence justifies splitting them as
biological (or recognition) species.
“To me the dilemma is whether we should
recognize 2 species (swainsoni and a
polytypic coronatus; Van´s Option B)
or 4 species (swainsoni, castelnaui, coronatus, and a polytypic mexicanus;
my new Option E). Both alternatives are compatible with phylogenetic data, and
the genetic divergence between the W Amazonian castelnaui and the mexicanus
group is nearly as deep as that between these two groups and coronatus. I examined the calls
unsystematically again, and the vocalizations seem very similar. It could be
that coronatus might have a slightly
higher pitched and perhaps shorter call than castelnaui, but the vocalizations are overall quite similar, and a
proper study is needed to increase our understanding of vocal variation in the
phylogroups uncovered by Reyes et al. 2023. Given the (apparent!) vocal
similarities and the not-so-deep (but also not-so-shallow) genetic differences
between castelnaui-coronatus-mexicanus,
I vote for option B for now. If new information appears, or someone with a
deeper understanding of vocalizations of the group uncovers diagnostic
differences in calls of these groups, I would be delighted to support my option
E, but at present I think that the information is not there to show that this
is the best option.”
Comments
from Naka:
“As you have guessed, there is a reason why we decided not to propose a
complete taxonomic rearrangement of this species in the Reyes et al. article,
mostly due to lack of i) nuclear data and ii) proper morphological and vocal
assessment in the clade. Although I agree with the major finding of the
article, that the six taxa involved likely represent six independent lineages,
I would not encourage, nor vote for a 6-species split at this stage. As we all
know, mDNA is great in reaching reciprocal monophyly,
yet it represents a bad proxy for gene flow. Therefore, we lack that key piece
of evidence to declare these taxa as fully independent lineages beyond a
reasonable doubt (meaning, these can fall apart in the next genomic study).
“I
personally like the idea of having three biogeographically-sound main lineages,
including i) a similar sounding Middle American/trans-Andean taxon (including
mexicanus, occidentalis, and fraterculus, although I wish the dry
forest occidentalis would sound any different!); ii) a rather distinct
Atlantic Forest taxon (swainsoni); and an Amazonian lineage (O.
coronatus + O. castelnaui). However, this arrangement contrasts with
the mtDNA results, which show that the two Amazonian forms are not sister taxa,
as first found by our study back in 2018 (Naka and Brumfield, 2018). Therefore,
the options here (if we are to follow the mtDNA tree), are to have 2, 4 or 6
lineages. At this stage, given the lack of better genetic resolution, and the
poor vocal sampling available, I will vote for option B.
In
short:
“A. NO, to Retain
traditionally defined broad O. coronatus.
“B. YES, to Recognize
two species (as in HBW and Birds of the World): O. swainsoni and O.
coronatus, which would include all other taxa.”
“C.
NO
“D.
NO”
Comments
from Stiles:
“A. YES; B. NO - the evidence for further splits is weak and inconsistent as it
stands: more data required: C. NO; D. NO.”
Comments
from Zimmer:
“A.
NO.
“B.
YES.
“C.
NO
“D.
NO.
“Personally,
the arrangement that makes the most sense to me intuitively, is the 4-way split
(i.e. Return to the classification of Cory & Hellmayr), but I don’t think
this option is rigorously defensible on current knowledge. I feel pretty confident from personal
experience that swainsoni is worthy of being elevated to species rank,
based upon noted vocal differences, genetic data, plumage, ecological
differences and distribution and how those fit established biogeographic
patterns. As an aside, I don’t think
that the illustrations included in the Proposal adequately convey just how
different occidentalis and swainsoni are in overall color from
the much duller Amazonian taxa that occupy the intervening areas – both of
those taxa, one occupying dry forest and the other Atlantic Forest are really
richly ochraceous and very different from other taxa in the complex. Regardless, I think the best course of action
for now, at least in terms of what is well supported by existing data, is the
conservative two-way split of swainsoni versus everything else. In that case, Van’s suggestion that we adopt
the existing BOW names of Atlantic Royal-Flycatcher for swainsoni, and
Tropical Royal-Flycatcher for everything else makes the most sense, with the
latter name being easily changed to adapt to any further splits.”
Comments
from Bonaccorso:
“
“A. NO. As several SACC members
have already acknowledged, there is preliminary evidence suggesting that
maintaining the traditionally defined O. coronatus may overlook the complexity of the group.
“Still,
obtaining complete vocal data for all populations could prove very difficult.
Based on our experience with the western Ecuadorian population (O. coronatus
occidentalis), gathering vocalizations from this taxon will be particularly
challenging due to its rarity, highly localized distribution, and potential
seasonal movements throughout the year.
“B. YES. For the time being,
based on limited nuclear (UCE) and mitochondrial (ND2) data (both topology and
distances), and qualitative vocal differences, it seems safe to call O. swainsoni a separate species from O. coronatus.
“C. NO. More data are
needed.
“D. NO. More data are
needed.”
Comments
from Glaucia Del-Rio (guest vote):
“A.
NO
“B.YES
“C.
NO
“D.
NO
“I
do not think the available data justify splitting beyond the two-species
treatment. As mentioned before, mitochondrial DNA, while useful for revealing
genealogical structure, is not sufficient to delimit species. For me, the
consistent placement of O. swainsoni as the deepest lineage in both
mtDNA and UCE studies, and its distinctive plumage and vocal differences,
provide sufficient evidence to recognize it a different species from the O.
coronatus complex.”