Proposal (1056) to South American Classification Committee
Treat the musculus
subspecies group as a separate species from Troglodytes aedon
Note
from Remsen: This is a NACC proposal that is here relayed
to SACC but severely pruned from the original NACC proposal to eliminate the
sections on all the extralimital Caribbean taxa (mostly now treated as separate
species by NACC); Brian Sullivan and I were co-authors on the original, but
that was strictly on the extralimital Caribbean taxa, which we predicted
therein were not a monophyletic group, subsequently confirmed by Klicka et al.
(2023). The NACC proposal passed 9-1,
and it also passed WGAC.
Effect
on SACC: This
would change what we currently consider as House Wren (Troglodytes aedon)
to Southern House Wren (T. musculus).
Background:
This
is an update of NACC Proposal 2022-B-10 by Remsen, Jaramillo, and Sullivan,
which proposed to recognize as many as seven species in the Troglodytes
aedon complex in the Caribbean. The proposal failed 5-6 (with 10c and 10g
failing 4-7). However, nearly all NACC members who voted no in 2022
acknowledged that multiple species must be involved in the complex, but that a
comprehensive integrative study is needed before action should be taken. Now, a
near-comprehensive phylogeny based on mtDNA and genomic data has appeared
(Klicka et al. 2023) and provides an opportunity to reevaluate the complex.
However, this now needs to be done from a broader perspective, given the
pattern shown in Klicka et al. (2023). In addition, reevaluation of other
papers provides a more integrative approach for examining species limits of all
but the rarest (or extinct) taxa. Although some of the relationships in the
complex clearly require further population and genetic sampling and analysis,
as indicated in Imfeld et al. (2024), we consider that sufficient data now
exists and is summarized herein to enable several changes to the current
taxonomy.
Committee
members can review the 2022 proposal (https://americanornithology.org/wp-content/uploads/2022/03/2022-B.pdf) and comments
(https://americanornithology.org/about/committees/nacc/current-prior-proposals/2022-proposals/comments-2022-b/#2022-B-10)
in conjunction with this proposal, as there is much information therein that is
not repeated here, mostly on the extralimital Caribbean taxa.
New
information:
More
taxa are included in the mtDNA phylogeny than in the genomic tree of Klicka et
al. (2023), and, not unexpectedly, the results differ somewhat. We focus here
on the genomic phylogeny and supplement that with data from the mtDNA phylogeny
for taxa missing from the former. Note
that Klicka et al. (2023) refer to all Caribbean samples as “T. a.
martinicensis”, even though the included samples are from Trinidad,
Grenada, St. Vincent, and Dominica, and thus must represent albicans, grenadensis,
musicus, and rufescens, respectively (this is made clear in the
supplementary materials). This may mislead some readers into thinking that one
taxon (martinicensis from Martinique) falls into multiple clades, which
is not the case; in fact, that extinct taxon is not included in the sampling.
Troglodytes
musculus.—The
most compelling result from the Klicka et al. (2023) phylogeny is that on
genomic data there are two main clades of Troglodytes aedon: the aedon
and musculus clades. The aedon clade (including brunneicollis)
occurs from southern Canada southward through the Isthmus of Tehuantepec, and
in Mexico it is largely a bird of pine-oak woodland in the highlands, whereas
the musculus clade occurs in a variety of habitat types and elevations
from southern Veracruz and northern Oaxaca through the Yucatán Peninsula and
southward through South America. The STRUCTURE results do not show
intergradation between these two groups, although sample size from the relevant
area is small.
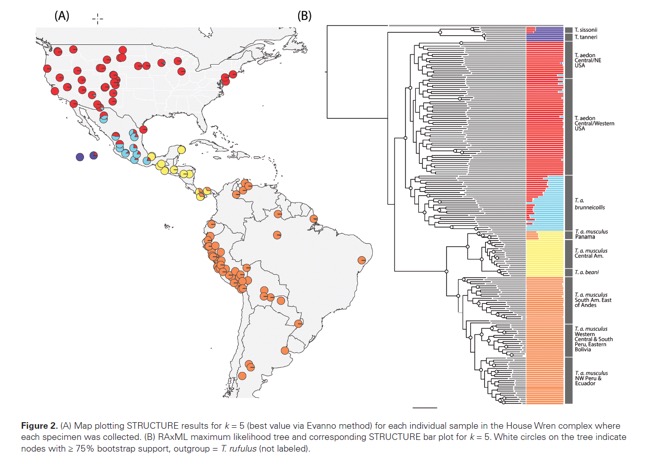
(from Klicka et al.
2023)
Howell
and Webb (1995) treated the musculus group separately, as “Troglodytes
aedon (in part) or T. musculus”. They considered the songs of these
two groups not reliably distinguishable, but Sosa-López and Mennill (2014)
found some vocal differences between them, as well as between other taxa that
are not presently considered candidates for species level.
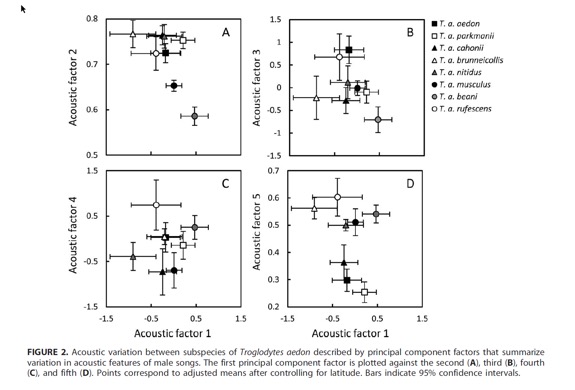
(from Sosa-López and
Mennill 2014)
These
authors also demonstrated mensural differences between the aedon and musculus
groups:
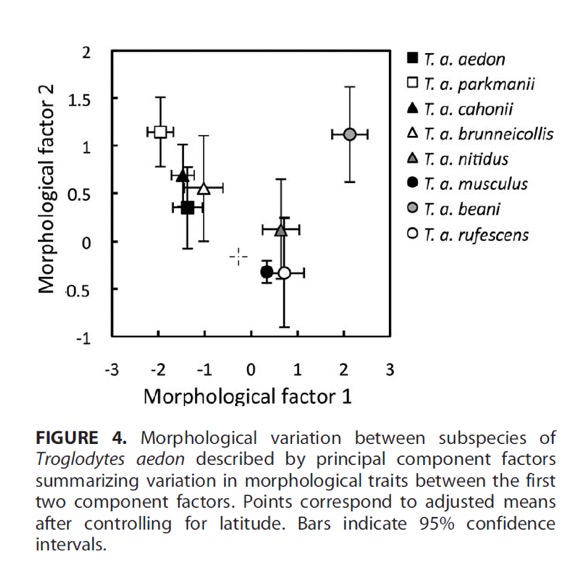 ’
’
(from Sosa-López and
Mennill 2014)
although
the disputed race nitidus (e.g., placed in the brunneicollis
group in Clements but not recognized at all by IOC-WBL) groups here with musculus,
not brunneicollis, etc. This seems unsurprising as nitidus is
from mountains of northern Oaxaca, thus near or at the contact zone between brunneicollis
and intermedius of the musculus group. But Nelson’s (1893)
description of nitidus indicates it is darker and more reddish-brown
than typical brunneicollis, so it could be variable, be more like brunneicollis
in color but more like intermedius in measurements, or perhaps both
forms occur in sympatry in the region. More study obviously needed on that.

(from Nelson 1893)
OD
of the geographically adjacent subspecies, intermedius Cabanis, 1861:

Google
translation:
“above murine-brown, wings and tail narrowly
barred with black; below with the eyebrows yellowish-brown; with hypochondria,
more and more diluting, blushing; with rufous subcaudal coverts, banded
transversely with black, interspersed with some whitish spots. This species is
similar to the americano and the platensi,
as if intermediate.”
“Just as brunneicollis is the
corresponding southern form of the more northern aedon [eastern Canada
and US], so intermedius could be viewed as that of the North American americanus
[=parkmanii] [western Canada and US]. At the
same time, our bird forms the transition from the northern species to those of
the South American continent through markings and coloring, as does its
geographical distribution. All northern species have the lower caudate coverts
vividly dark and brightly spotted, and this marking extends more or less over
the wings and anal area, while in almost all South American species the same
parts of the body are almost entirely without transverse markings. In intermedius
only the undertail coverts are clearly cross-banded. Our bird differs from americanus
in its darker upper side, which turns grey-brown on the crown and neck;
furthermore, by the light brownish color of the underside, which is reminiscent
of platensis Neuw., but is darker and less
pale. The back of platensis is also not so brown, but more of a
gray-brown color, and the transverse markings on the lower tail coverts are
less developed, almost only present as tip markings on the individual feathers.”
In
essence, Cabanis was stating that his new form intermedius is similar to
western North American birds, not to the geographically adjacent brunneicollis.
In any case, if there is a zone of intergradation, it seems it must be a narrow
one. And this marked discontinuity in phenotype in near-parapatry has been
known a long time and has now been demonstrated to be real based on the Klicka
et al. (2023) phylogeny, although a larger sample size in this region and more
focused study on this aspect is needed to better understand the interactions
between these two forms.
Sosa-López
et al. (2016) analyzed response to playback with respect to mtDNA divergence:
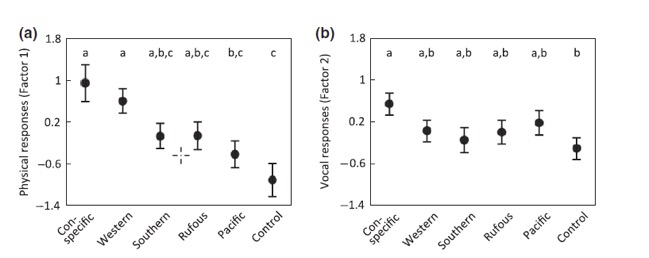
(from Sosa-López et al.
2016)
and
found that brunneicollis (a, b) responded more strongly physically (but
not vocally) to Western than to Southern house wrens, with the response to the
latter at the level of the non-conspecific Rufous-browed and only slightly
higher than to Pacific Wren.
So,
there is a good case to be made for splitting the intermedius group of musculus
from the aedon group including brunneicollis. The most obvious
problem with this is that there is some gene flow in Panama with the South
American musculus group, so simply splitting musculus from the
southern Mexico-Central American intermedius group requires further
study and has nomenclatural implications. And, as for the evidence for
considering the intermedius and musculus groups separate species,
if it has ever been seriously considered it is not evident in the papers
reviewed here, and for now we consider intermedius a group within the musculus
complex.
Troglodytes
brunneicollis.—mtDNA
phylogenies including that in Klicka et al. (2023) showed brunneicollis
as paraphyletic with several outgroups, thus strongly suggesting species status
for brunneicollis. However, this result has not held up in the genomic
analysis of Klicka et al. (2023), and furthermore broad introgression with aedon
in the southwestern USA has been shown to occur (as was already apparent from
plumage).
Troglodytes
parkmanii.—As with brunneicollis, mtDNA
phylogenies, including that in Klicka et al. (2023), showed parkmanii
as paraphyletic, but this is again not the case for the genomic phylogeny, in
which it groups with aedon. They have been shown to be vocally different
and to have some reduced response, but the differences are not at the scale of
the other taxa studied (for which see below).
Recommendations: (The original proposal included separate
recommendations, both NO, on whether to split brunneicollis from aedon,
and whether to split parkmanii from aedon;
both were voted down, and given that these extralimital taxa are in the NACC
area only, that part of the proposal is excluded from the SACC version).
We
strongly recommend a YES vote to splitting musculus (including the intermedia
group) from the aedon group (including the brunneicollis group).
Should the intermedia group be split later from the musculus
group, the species-level nomenclature would have to change for the Central
American group to intermedia, but we think the evidence for the split
between the aedon and musculus groups is overwhelming.
English
names:
NACC went with Northern House Wren and Southern House Wren. Unless someone objects and would like to do a
separate proposal on English names, I think we should follow NACC. Note the absence of a hyphen because the two
are not necessarily sister taxa with respect to Caribbean taxa or T. cobbi.
Literature
Cited (in
the original, lengthier proposal):
Cyr, M.È., K. Wetten, M.H. Warrington, and N. Koper. 2020. Variation in
song structure of house wrens living in urban and rural areas in a Caribbean
small island developing state, Bioacoustics https://doi.org/10.1080/09524622.2020.1835538
del Hoyo, J., and N.J.
Collar. 2016. HBW and BirdLife International Illustrated Checklist of the
Birds of the World. Volume 2: Passerines. Lynx Edicions, Barcelona.
Imfeld, T.S., F.K.
Barker, H. Vázquez-Miranda, J.A. Chaves, P. Escalante, G.M. Spellman, and J.
Klicka. 2024. Diversification and dispersal in the Americas revealed by new
phylogenies of the wrens and allies (Passeriformes: Certhioidea). Ornithology
(early view).
Kirwan, G.M., A.
Levesque, M. Oberle, and C.J. Sharpe. 2019 Birds of the West Indies. Lynx and
BirdLife International Field Guides. Lynx Edicions, Barcelona.
Klicka, J., K. Epperly,
B.T. Smith, G.M. Spellman, J.A. Chaves, P. Escalante, C.C. Witt, R.
Canales-del-Castillo, and R.M. Zink. 2023. Lineage diversity in a widely
distributed New World passerine bird, the House Wren. Ornithology 140:1-13.
Sosa-López, J.R., and
D.J. Mennill. 2014. Continent-wide patterns of divergence in acoustic and
morphological traits in the House Wren species complex. The Auk: Ornithological
Advances 131: 41-54.
Sosa-López, J.R., J.E.
Martínez Gómez, and D.J. Mennill. 2016. Divergence in mating signals correlates
with genetic distance and behavioural responses to
playback. Journal of Evolutionary Biology 29:306-318.
Wetten, K.N. 2021.
Morphological divergence in the House Wren (Troglodytes aedon) species complex:
A study of island populations with a focus on the Grenada House Wren (T. a.
grenadensis). Master’s Thesis, University of Manitoba.
Pamela C. Rasmussen,
May 2025
___________________________________________________________________________________________________
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Claramunt:
“YES. Both mitochondrial and nuclear-genomic data indicate the existence of two
independent lineages that, despite geographic proximity, have not been
exchanging genes. Response to each other’s songs is moderate, but there must be
other mechanisms of isolation in order to prevent gene flow, which is not
happening.”
Comments from Areta: “YES. This is a solid result supported by differences in plumage,
habitat, vocalizations, and genetics.”
Comments
from Lane:
“NO. Honestly, I am having a bit of a hard time navigating the proposal and
feeling like the conclusions are as solid as they are made out to be. My main
issue is that the crux of the problem is in the Gulf slope of Mexico north/west
of the Isthmus of Tehuantepec, where the brunneicollis group (here a
member of the northern aedon group) and the intermedius group
(here a member of the musculus group) come into contact or are
parapatric. To me, this is the only important area, and it is hardly addressed
with more than a little lip service. I am assuming that the Klicka et al.
(2023) paper is the primary source for the phylogenetics used in the proposal,
but in that paper, there are not many samples made from the crucial area of the
lowlands of Oaxaca and the nearby highlands. There is mention made of the named
taxon nitidus, but then it is effectively ignored even though it was
described, then synonymized by Binford (1989) with brunneicollis. The
proposal mentions that nitidus is a musculus population, but I
don't understand the basis for that statement! Klicka et al. don’t seem to have
sampled that population, and phenotypically it appears to be very similar to brunneicollis
(as per Binford’s comment), so how was it determined to be a member of the musculus
group? In the supporting files of Klicka et al, I see 3 Veracruz samples
(presumed intermedius) and 3 Oaxaca samples (presumed brunneicollis).
In the mtDNA tree built there (and reflected in Figure 1 of the main paper), it
is noteworthy that the bulk of the brunneicollis clade is not only not
sister or embedded within the aedon clade, but outside the entire T.
solstitialis clade as well! However, the six samples from the
Veracruz/Oaxaca area are not found to fall out together, nor do they seem to in
the SNP tree that is reflected in Figure 2 of the paper (and reproduced in the
proposal). But this small sample does not quench my interest in what exactly is
going on at this crucial place! I would really like to see a more in-depth
study of the lowland Gulf slope populations and the highland brunneicollis
here to determine if they truly are not interbreeding. Do they even have the
opportunity to interact and remain elevationally/habitatly
(did I just invent a word?) parapatric? In my limited experience in the area,
they seem to be parapatric. But overall, I am remarkably underwhelmed by any
phenotypic and vocal differences between the musculus group and aedon/brunneicollis
group(s). In fact, to me brunneicollis is by far the most distinctive in
voice and habitat choice of the whole complex! So, to have spent so little
effort in trying to investigate this limited area of potential interaction in
eastern Mexico makes me nervous in drawing such large conclusions about the
complex. Until I feel like that is done, I just can’t bring myself to agree
with this split. NO.”
Comments
from Stiles:
“YES to splitting of musculus from aedon; I note that Dan´s
worries about nomenclature are all within the aedon clade and don't
alter the split of musculus.”
Comments
from Remsen:
“YES, but with extreme reluctance. If
NACC and AviList had not already endorsed the split and if the 2-species
treatment had been novel instead of a widespread treatment in many earlier
classifications, I would vote NO. I
share Dan’s concerns – this all makes me uneasy because despite all of the new
data on genetics and voice, no one has critically studied the contact zone in
Mexico. Contact zones provide direct evidence
on how populations interact with each other in terms of gene flow, and they remove
the need to make inferences and indirect yardstick comparisons. My YES vote is based on the assumption that,
as noted in the proposal, that if the two populations intergrade in Mexico,
then this would have been known a long time ago from studies of specimens, so
even though the genetic sampling in the critical area is remarkably weak in the
new studies (as noted in Pam’s proposal), all signs, phenotypic or genotypic
(the deep genetic split between the clades), point to an absence of a hybrid
swarm. Binford, for example, would very
likely have picked up on this in his studies of the avifauna in Oaxaca. So, if forced to justify my vote based on
more than momentum and the palliative of returning to a previous taxonomy, this
absence of any sign of a hybrid swarm would be my main argument. Regardless, what’s going on in that contact
zone would be of general interest and, as far as I can tell, would be an easy
field study.
“As far as the evidence discussed in
the papers as outlined in the proposal, much of it, albeit interesting and
important, is not particularly relevant to species limits. The differences in measurements and plumage are
not really relevant to species limits unless used as a proxy for gene flow in
the vicinity of the contact zone.
Similarly, habitat differences in this complex are about as
irrelevant as they can be. Just within
the musculus group in Bolivia, one can find House Wrens singing in
frigid dusty towns in the Titicaca Basin to sweltering jungle towns in the
Beni, thus spanning a remarkable climatological range. The North American aedon group spans
an ecological gradient almost as steep, from aspen groves at 10,000 feet in
Colorado to near sea level in hot humid coastal North Carolina. When we invoke “integrative” taxonomy in our
decisions, there is a tendency to ignore the point that many of the points that
are integrated are irrelevant to species limits unless they apply to contact
zone regions and thus acts as a proxy for gene flow. I think we take too much comfort in listing a
bunch of phenotypic differences that are not directly relevant to species
limits unless from regions in which there are contact zones.
“As for the playback experiments, if we
take them as evidence for splitting musculus from aedon, then we
should also take them as evidence for splitting brunneicollis from aedon,
as advocated by Sosa-López and Mennill (although that is out of SACC’s
geographical purview), despite the Structure plot that shows some evidence of
gene flow. In general, playback
experiments in wrens seem tricky, as evidenced by the Sosa-López results. Anecdotally, for example, here in Louisiana
in winter, House, Sedge, Marsh, and Carolina will regularly respond moderately
aggressively to playback of each other’s songs.
“So, after all that preaching, I
conclude no real harm done in returning to species limits that were in place in
many earlier classifications despite absence on nail-in-coffin study of contact
zones.”
Comments
from Robbins:
“I too feel on the fence on this one given known hybridization between birds in
central Panama and the lack of adequate sampling of the two genetically
distinct clades at the zone of contact in southeastern Mexico. I don't have
access to specimen material to see if birds involved in hybridization in
central Panama are less or more distinct than birds in contact in SE
Mexico. Having said that, we know plumage in at least North American
wrens has little to do with species limits, a good example is Eastern and
Western Marsh Wrens — virtually identical in plumage but have distinct songs
and are quite different genetically.
“Ascertaining
relevant differences in vocalizations is difficult in this entire
complex. Within migratory US populations, broadly speaking, western birds
sound distinct from eastern birds, so are relatively minor differences in
vocalizations in other areas meaningful? I suspect one would find song
variation among all groups that are not migratory. I consider playback
experiments to be of little value with this group, as Van correctly points out,
one can get intergeneric response in at least North American wrens.
“So,
in my opinion, we are basing this decision on genetics without enough
information on the contact zone to help guide what is the best course of
action. If NACC hadn't voted for the split, I would vote NO and would wait
until we have more information from the contact zone in SE Mexico. But, given
their decision, with reservations, I'll vote YES for recognition of the split.”
Comments
from Bonaccorso:
“YES. The deep split between these two taxa seems to be telling a lot
about the historical separation of these two lineages. Even if there is not
enough sampling from a potential contact zone, the lack of Central America
genotypes in the North American Lineage and vice-versa seems to indicate that
there is no current gene flow. Also, even if there was hybridization at the
contact zone, if genetic distance is any indication of potential reproductive
incompatibilities, it is likely that such hybridization is very restricted and
constantly limited by natural selection.”
Comments
from Naka:
“YES. I have been using musculus for the South American populations for a very
long time now, and I am happy to maintain that treatment in face of the strong
molecular evidence showing that they represent independent lineages with no
signs of admixture (or historical gene flow) between any of the musculus
and aedon populations. Also, I consider that most of the discussions
about gene flow between brunneicollis and nominate aedon, do not
affect our treatment for a South American musculus, which is strongly
supported by molecular data. I agree that vocal and phenotypic data may not be
that important in a species with such high climatic and habitat plasticity.
Now, If you ask me what I think would happen if we throw a population of musculus
in Mexico city or Texas, I would remain silent. But I
do believe that this has not happened for a reason and suspect the advance of aedon
into the south or musculus further north was likely prevented by
competitive exclusion.”
Comments
from Zimmer:
“YES. The evidence for two main clades on independent evolutionary trajectories
appears strong, even if the contact zone in Mexico hasn’t been well sampled
from a genetic standpoint. As Van points
out, if there was a broad hybrid zone in southern Mexico, it almost surely
would have been already known from examination of specimens. I’ve never been overly impressed with vocal
differences in this complex (other than those between insular Caribbean taxa
versus mainland taxa, which are VERY different), but that having been said, I
would agree with Mark and Van that voice and response to playback are probably
not nearly as meaningful with wrens in general as they are in other
groups. Similarly, I think habitat is
incredibly plastic within this group, and really shouldn’t factor into the
equation. I share Dan’s queasiness with all
of the questions concerning the true affinities of intermedius and with
the lack of data from contact zones in southern Mexico in general, but I think,
in the absence of strong evidence of intergradation/hybridization, the genomic
and mtDNA analyses referenced in the Proposal should be considered sufficient
to recognize the proposed split of the musculus-group from the aedon-group.”