Proposal (1057) to South
American Classification Committee
Revise
species limits in Automolus ochrolaemus.
A. Treat extralimital Central American exsertus as a separate
species; B. Treat extralimital Middle American cervinigularis as a
separate species (from exsertus and ochrolaemus)
Note
from Remsen:
The two parts of this proposal are separate proposals relayed from NACC, which
treated exsertus as a separate species in 2018 (Chesser et al. 59th
Supplement 2018; proposal 2018-A-2) and cervinigularis as a separate
species in 2024 (Chesser et al. 65th Supplement 2024; proposal 2024-c-22;
passed 8-2). We have to process these
because they affect what SACC considers to be A. ochrolaemus and also
because these extralimital splits potentially affect our English names.
Also,
the sequence of the proposals and the names can be confusing because they were
separate proposals in NACC. The first
one A deals with splitting Pacific slope exsertus from everything else,
namely broadly defined ochrolaemus, and then B deals with splitting the
rest of the Middle American populations (as cervinigularis) from the
basically South American ochrolaemus.
Effect
on SACC:
No effect other than to change what we would assign to the range of A.
ochrolaemus
Our
current SACC note reads:
85c. Chesser at al. (2018)
treated the Central American exsertus group as a separate species
following the results of Freeman and Montgomery (2017). Dyer and Howell (2023) treated exsertus
group and the Middle American cervinigularis group as separate species
from A. ochrolaemus. Chesser et
al. (2024) treated Middle American cervinigularis as a separate species
from A. ochrolaemus based on comparative genetic distances (Smith et al.
2014). SACC
proposal needed.
Part
A. Treat Automolus ochrolaemus exsertus as a separate species from Buff-throated
Foliage-gleaner A. ochrolaemus
Background: Automolus ochrolaemus, the Buff-throated
Foliage-gleaner, is found in humid lowland forests across a large swath of
Central and South America. There is substantial plumage, vocal and genetic
variation within its broad distribution (see Smith et al. 2014 for
phylogeographic structure, and Remsen 2017 for descriptions of plumage and
vocal variation). This proposal concerns two of the populations found within
Central America: exsertus, which
occurs on the Pacific slope of Costa Rica (and adjacent western Panama), and hypophaeus, which is found along the
Caribbean slope of Central America. These two populations are geographically
isolated and ~ 6% different in mtDNA (cyt b), suggesting they last shared a
common ancestor around 3 million years ago. They differ in voice and are roughly
similar in plumage (Remsen 2017).
New information: Freeman and Montgomery (2017) conducted playback
experiments on 15 territories of exsertus
and 14 territories of hypophaeus.
Each playback experiment measured whether populations discriminated against
song from the other population; thus, these experiments simulated secondary
contact between these two geographically isolated populations. Briefly, each
experiment measured the behavioral response of a territorial bird to two
treatments: 1) song from the local population (sympatric treatment) and 2) song
from the allopatric population (allopatric treatment). All territorial birds
responded to sympatric song by approaching the speaker (typically to within 5
m).
We defined song discrimination as
instances in which the territory owner(s) ignored allopatric song, defined as a
failure to approach within 15 m of the speaker in response to the allopatric
treatment. We calculated song discrimination for each taxon pair as the
percentage of territories that failed to approach the speaker in response to
allopatric song. For example, a song discrimination score of 0.8 indicates that
80% of territorial birds (e.g. 8 out of 10) ignored allopatric song while
simultaneously actively defending a territory. We assume that song
discrimination is a proxy for premating reproductive isolation; that is, our
experiments provide insight into whether these populations would recognize each
other as conspecific and interbreed (or not) were they to come into contact
with one another. It is unknown what degree of song discrimination is “enough”
that song constitutes a strong enough premating barrier to reproduction that
allopatric populations merit classification as distinct biological species. To
provide a yardstick, we considered nine allopatric Neotropical taxon pairs that
were recently split (or have pending proposals to the South American
Classification Committee) in part based on differences in vocalizations. We
found the average song discrimination in these nine taxon pairs to be ~ 0.6
(60% of territorial birds ignored allopatric song) and suggest that species
limits deserve to be reconsidered when taxon pairs currently classified as
subspecies have song discrimination scores above ~ 0.6.
We found that 13 out of 15 territorial
birds from the Pacific slope of Costa Rica (exsertus)
discriminated against song playback of Caribbean hypophaeus (discrimination = fail to approach within 15 m of the
speaker). Results were similar in the
opposite direction: 12 out of 14 territorial birds from the Caribbean slope of
Costa Rica (hypophaeus) discriminated
against song playback of Pacific exsertus
(discrimination = fail to approach within 15 m of the speaker).
Recommendation:
Populations of Buff-throated
Foliage-gleaners from the Pacific and Caribbean slopes of Costa Rica respond
strongly to local song but essentially ignore song from their relatives across
the mountains. This suggests that vocal differences constitute a strong
premating barrier to reproduction between these taxa and is consistent with the
genetic data that indicates that, despite their close geographic proximity,
these populations last shared a common ancestor ~ 3 million years ago. I
therefore recommend treating exsertus and
hypophaeus as distinct biological
species. In practice, this means that I recommend treating exsertus as a distinct biological species from the entire rest of
the Buff-throated Foliage-gleaner complex. There may be additional biological
species lurking within this complex. For example, we documented strong song
discrimination between hypophaeus and
pallidigularis (found in eastern
Panama and northwestern South America; these two taxa presumably interact in a
contact zone in central Panama and may prove to be distinct biological species.
As for an English name for exsertus,
Ridgway (1911) and Cory and Hellmayr (1927) called it "Chiriqui
Automolus", which would translate to "Chiriqui Foliage-gleaner",
which would seem to be a reasonable name.
Literature
Cited:
Freeman,
B. G., & Montgomery, G. A. 2017. Using song playback experiments to measure
species recognition between geographically isolated populations: A comparison
with acoustic trait analyses. The Auk 134(4), 857-870.
Remsen,
J.V., Jr (2017). Buff-throated Foliage-gleaner (Automolus ochrolaemus).
In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A. & de Juana, E.
(eds.). Handbook of the Birds of the World Alive. Lynx Edicions,
Barcelona. (retrieved from http://www.hbw.com/node/56571 on 20 October
2017).
Smith,
B.T., McCormack, J.E., Cuervo, A.M., Hickerson, M.J., Aleixo, A., Cadena, C.D.,
Perez-Eman, J., Burney, C.W., Xie, X., Harvey, M.G. and Faircloth, B.C., 2014.
The drivers of tropical speciation. Nature 515.
406-406.
Benjamin
Freeman, May 2025
Part
B. Treat Automolus cervinigularis as a separate
species from Buff-throated Foliage-gleaner
A. ochrolaemus
Description of the
problem: Automolus ochrolaemus (Tschudi, 1844)
is a wide-ranging polytypic species of the lowland Neotropics, found in lowland
rainforest nearly throughout the Neotropics from southern Mexico through the
Amazon Basin. Until 2018, this was generally considered a single polytypic
species with seven subspecies. From north to south, these taxa are: cervinigularis (Sclater, 1857) of Mexico
to Guatemala; hypophaeus Ridgway,
1909 of Nicaragua to northwestern Panama on the Caribbean slope; exsertus Bangs, 1901 of the Pacific
slope of Costa Rica and far southwestern Panama; pallidigularis Lawrence, 1862 of eastern Panama south through the
Choco to northwestern Ecuador; turdinus
(Pelzeln, 1859) of the western Amazon Basin and Guiana Shield; ochrolaemus (Tschudi, 1844) of the
southwestern Amazon Basin; and auricularis Zimmer, 1935 of the southeastern
Amazon Basin (Birds of the World, 2023). Another subspecies, amusos, is sometimes recognized from
Honduras and Nicaragua, or considered a synonym of cervinigularis or hypophaeus.
Ridgway (1911) considered the complex to
comprise three species: A. cervinigularis
(including hypophaeus), A. pallidigularis (including exsertus), and although not covered in
his volumes, implicitly considered A.
ochrolaemus for all the extralimital taxa.
NACC proposal 2018-A-2 elevated exsertus to species rank based on
allopatry from hypophaeus,
mitochondrial divergence, and song discrimination in playback trials between exsertus and hypophaeus, and NACC adopted the English name Chiriqui
Foliage-gleaner. NACC adopted this split 8-2, with some committee member
comments mentioning that the rest of the taxa found west of the Andes might
eventually be split from the Amazonian taxa, pending further research. WGAC, in
addressing discrepancies among global lists, also chose to adopt the split of A. exsertus, but went one step further
and split the other two Middle American taxa as a species (A. cervinigularis, with hypophaeus)
separate from the South American taxa. They opted, however, to retain pallidigularis of eastern Panama and the
Choco with A. ochrolaemus of the
Amazon Basin and Guiana Shield.
New information:
1. Genetics:
Smith et al. (2014) sampled all taxa in this
group using the mitochondrial gene cytochrome b and estimated a gene tree that
was fairly well resolved. Below is the supplemental figure for the genus from
Smith et al. (2014) showing the sampling map, ecological niche model, and the
gene tree. On the left is the time-calibrated gene tree, and on the right is
the same tree with species as circumscribed by the species delimitation method
bGMYC.
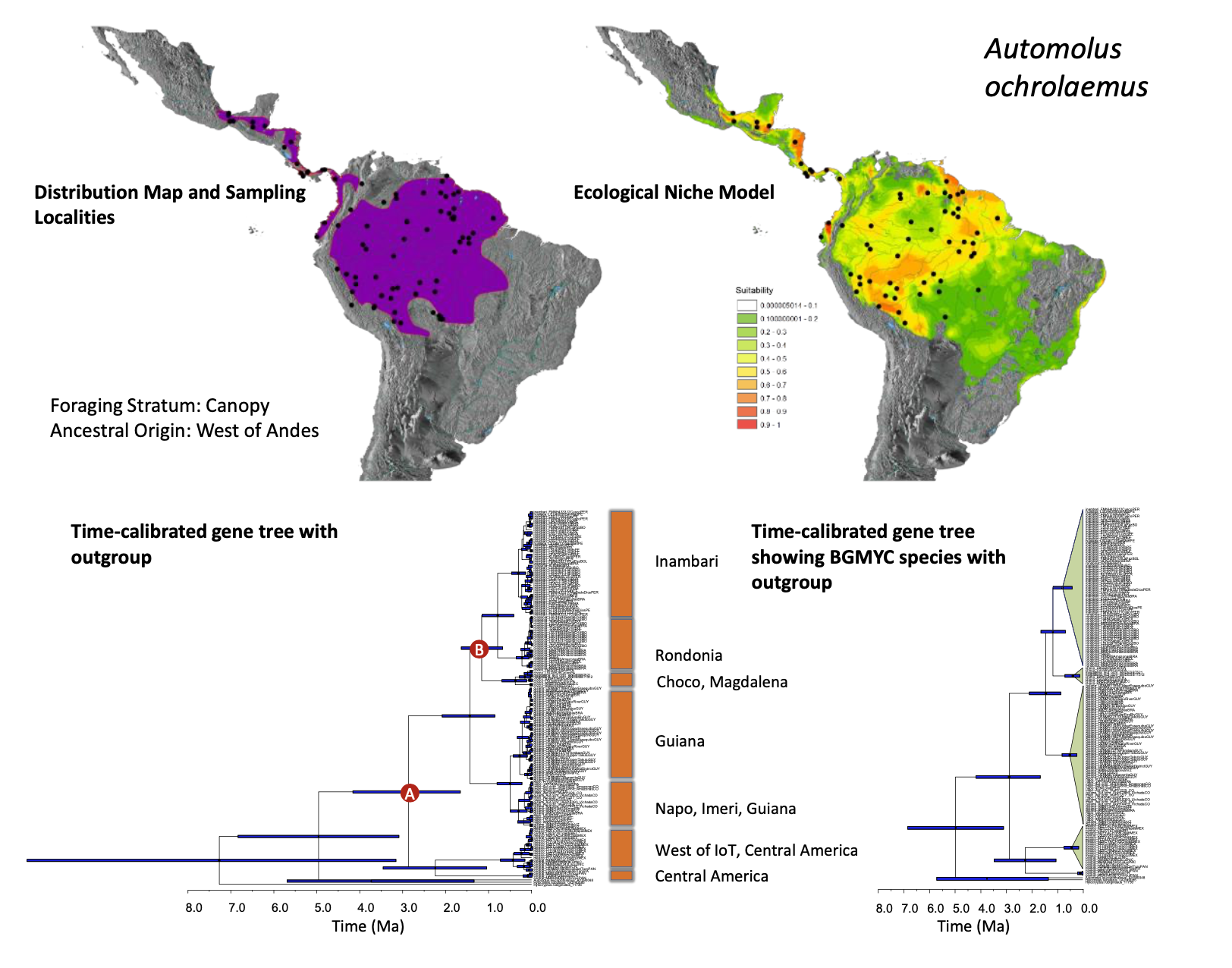

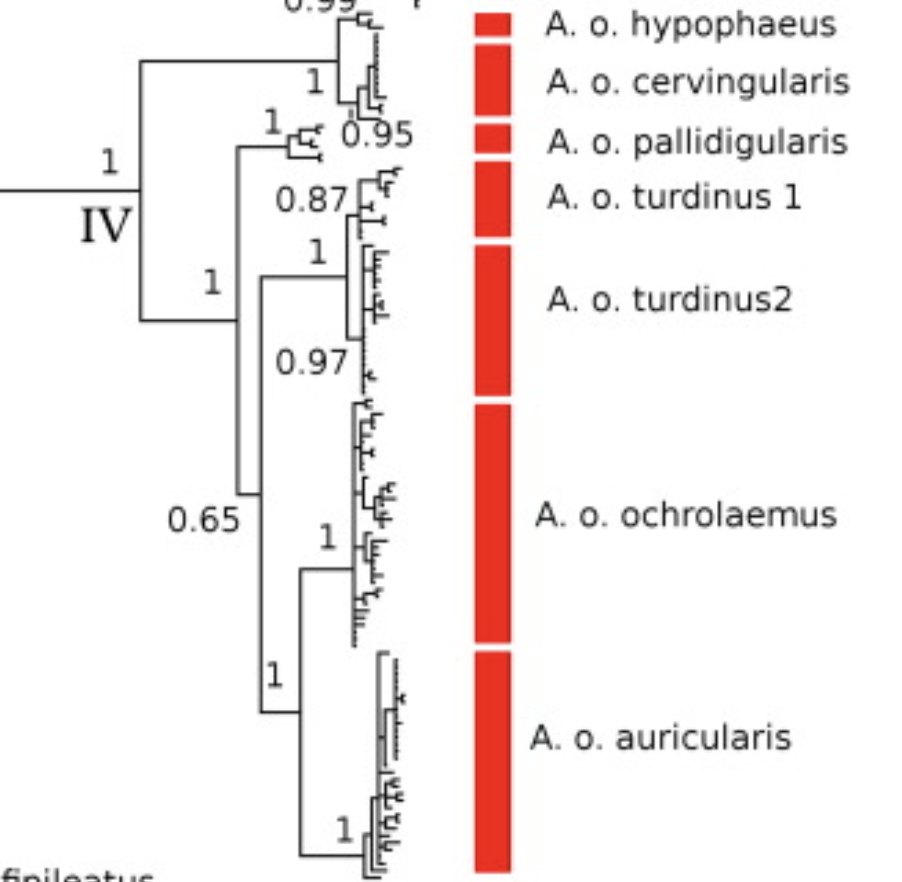
Broadly, Smith et al. (2014) found that exsertus and cervinigularis/hypophaeus were sister groups, which in turn were
sister to the remainder of the ochrolaemus
complex. The Choco taxon pallidigularis was
embedded within the rest of the South American taxa from east of the Andes. To
better illustrate these relationships, I have included an enlarged version of
the bGMYC tree above, with the corresponding taxa labeled to the right of each
cluster. Note that that outgroup has been removed here to better highlight the
few samples of exsertus at the bottom
of the tree. The time scale is in millions of years.
Shultz et al. (2017) sampled two mitochondrial
genes (ND2 and cytochrome b) for most taxa in the group, except exsertus, and recovered a similar
topology to Smith et al. (2014). However, they recovered pallidigularis as sister to the remaining South American taxa
rather than embedded within them. Shultz et al. (2017) also sequenced three
nuclear genes for these samples, but they showed no differentiation across the
group, as is often the case. They are not shown here.
Harvey et al. (2020) sampled two individuals in
the complex: one hypophaeus from
Costa Rica and one turdinus from
Peru. These samples were sisters, with a divergence time of about 2 Ma, which
is consistent with the mitochondrial data, but doesn’t provide much information
for taxonomy.
Claramunt et al. (2013) sequenced three
mitochondrial and three nuclear genes and recovered a topology consistent with
the other studies included here, with pallidigularis
clustering with the Amazonian taxa, and hypophaeus
sister to the rest (exsertus, cervinigularis, and auricularis not sampled). A portion of the tree from Claramunt et
al. (2013) is shown below.

2. Voice:
Although nothing is published on vocalizations
that I can find, there is considerable vocal variation in the group, and this
is partly the basis for elevating exsertus
to species rank. In listening to vocalizations, there are essentially two vocal
groups in the complex: one with a slower series of nasal descending notes
comprised of ochrolaemus, auricularis, turdinus, and pallidigularis,
and another with a faster song comprised of harsher notes sometimes strung into
a longer rattle. This latter group is comprised of cervinigularis, hypophaeus,
and (to an extent) exsertus. In
listening to recordings, it does seem like exsertus
(https://birdsoftheworld.org/bow/species/butfog4/cur/multimedia?media=audio) has consistently slower songs than cervinigularis/hypophaeus (https://birdsoftheworld.org/bow/species/butfog9/cur/multimedia?media=audio) with a somewhat harsher quality to the notes (thus, very unlike the
South American group). Some recordings of cervinigularis/hypophaeus also have a two-parted aspect
to the song, with the note shape being distinctly different in the first half
of the song, and with the second half often trailing off into a long rattle.
Songs of the three Amazonian taxa (ochrolaemus, auricularis, and turdinus)
are remarkably constant across their range: https://birdsoftheworld.org/bow/species/btfgle1/cur/multimedia?media=audio. However, pallidigularis adds
a bit of complexity to the matter. Recordings from the southern end of its
range in Ecuador closely resemble those of the Amazonian taxa (e.g. https://macaulaylibrary.org/asset/140267791, whereas those at the
northern end of its range in eastern Panama and northwestern Colombia are a bit
faster (https://macaulaylibrary.org/asset/60369, https://macaulaylibrary.org/asset/286906, https://macaulaylibrary.org/asset/610186681). That variation aside, there does seem to be a rather sharp break
between sweeter-sounding birds in the Canal Zone (e.g. https://macaulaylibrary.org/asset/28412) and rattling birds in the far west of Panama on the Caribbean slope (https://macaulaylibrary.org/asset/214530601). It is also possible that some of the recordings linked above are
after playback so the birds may be singing a more intense song, as at least
some recordings from the Canal Zone are slower and much more like Amazonian
birds (https://xeno-canto.org/448169).
Plumage:
David Vander Pluym was nice enough to photograph
a series of specimens of the taxa in this complex that are housed at the
Louisiana State University Museum of Natural Science (LSUMNS). Photos in
dorsal, lateral, and ventral views are shown below. In each photo, 2-3
individuals of each taxon are shown, with the taxon name written above, and the
red vertical lines separating the proposed species. The taxa from left to right
are: auricularis, ochrolaemus, turdinus, pallidigularis,
exsertus, hypophaeus, and cervinigularis.



Ridgway (1911) had some insights into the
plumage and morphometrics of this complex. As is noted above, he recognized
three species in the group, although these are not the arrangement that is
currently being considered, it is also not the broad polytypic circumscription.
Regarding cervinigularis, he noted:
“Mexican specimens
average decidedly deeper in color than others, especially the buff of
superciliary stripe, throat, etc., and brown of pileum, the latter almost sooty
in its darkness. Guatemalan examples have the back, etc., more rufescent or
castaneous, those from Honduras, British Honduras, and Nicaragua more
olivaceous than Mexican specimens. The series examined is, however, inadequate.”
For hypophaeus, he noted that it was “Similar
to A. c. cervinigularis but
coloration decidedly darker, especially under parts of the body, which are
isabella color medially darkening laterally into deep buffy olive, contrasting
strongly and abruptly with the buff or ochraceous-buff of chin and throat.” Ridgway (1911) considered specimens from Veraguas in western Panama
to be hypophaeus but had no samples
between there and the Canal Zone, which he considered pallidigularis. Thus, no specimens from the potential contact zone
were available to him. Regarding these samples of pallidigularis, he noted that it was “Somewhat
like cervinigularis but superciliary
stripe much less distinct (the supra-auricular portion more or less obsolete),
general coloration paler, feathers of chest without darker margins, and size
smaller.” Regarding exsertus,
which he considered a subspecies of A.
pallidigularis, he said it was “Similar to A. p. pallidigularis but slightly
larger, with relatively longer bill, color of back, etc., more olivaceous,
chest uniform in color, and buff of throat, etc., deeper.”
All of Ridgway’s comments seem (unsurprisingly)
consistent with the patterns shown in the photos above. Plumage is clearly
conserved across the group, with more intra-specific (if the split is adopted)
than inter-specific plumage variation. To my eye, the pale throat of pallidigularis really stands out, as do
the generally warmer underparts of cervinigularis.
However, other characters like bill size, degree of mottling on the chest, and
intensity of the olive below, all seem to vary.
Cory and Hellmayr (1925) merged all taxa into a
polytypic A. ochrolaemus, which seems
to be the basis for much of the modern treatment of the group until exsertus was elevated to species rank by
NACC. Although they did not elaborate on the decision to merge all these taxa
into one species, the following footnote for pallidigularis is of particular interest:
“It will be remembered that Salvin and Godman (Biol. Centr.-Americ,
Aves, 2, p. 158, 159) record both A.
cervinigularis and A.
"pallidigularis" from the Veraguas. Although no specimens are
available I have little doubt that all the birds of that region will ultimately
prove to belong to A. o. exsertus.
One of our Bogava skins, by reason of its distinct postocular stripe and
decidedly rufous under tail-coverts, closely approaches the eastern hypophaeus, and it is probable that
similar examples (which obviously represent only the extreme of individual
variation) have given rise to the reported occurrence of “cervinigularis" in the Veraguas.”
Bogava is in the
lowlands of Chiriquí, well within the range of exsertus. If I am interpreting this passage correctly, Cory and
Hellmayr (1925) are suggesting that exsertus
from the far east of its range might approach hypophaeus in plumage. There are low passes over the Talamancas in
this region, and it is possible that the two taxa might locally be in secondary
contact.
Recommendation: I very tentatively recommend a YES vote on splitting cervinigularis from ochrolaemus, which would bring us in line with WGAC. The vocal
differences between the cervinigularis
group and the ochrolaemus group are
certainly much greater than those that led to the split of exsertus from cervinigularis.
However, those taxa are allopatric and had good data on vocal discrimination,
whereas cervinigularis and ochrolaemus are certainly in contact in
west-central Panama but without any data from the contact zone.
The split then rests on two primary factors: the
primarily mitochondrial gene trees showing a close relationship between cervinigularis and exsertus (thus rendering ochrolaemus
sensu lato paraphyletic if it
includes the cervinigularis group)
and the very qualitative assessment of vocal differences included here. Those
two factors are admittedly quite highly differentiated, and more so than are
the differences that led to the split of exsertus.
WGAC used this same reasoning to strongly advocate for the split of cervinigularis.
WGAC adopted the English common names of
Fawn-throated Foliage-gleaner for A.
cervinigularis and Ochre-throated Foliage-gleaner for A. ochrolaemus, and if the split passes I recommend that we follow
suit. Because this is a parent-daughter split (mostly, anyway), new names
should be adopted for the daughter species and Buff-throated should be
abandoned. I note, however, that Ridgway (1911) used Buff-throated for cervinigularis sensu stricto. ‘Cervinus’ refers to ‘stag-like’, hence the common
name of Fawn-throated, which I think is a good name. Ochre-throated also
parallels the scientific name for ochrolaemus
and is similar to the “Ochraceous-throated” used by Cory and Hellmayr (1925).
Ridgway (1911) used Pale-throated for pallidigularis
sensu stricto, which is an
appropriate name if that taxon is eventually elevated to species rank.
Literature Cited:
Birds of the World. 2023. Edited by S. M. Billerman, B. K. Keeney, P. G.
Rodewald, and T. S. Schulenberg. Cornell Laboratory of Ornithology, Ithaca, NY,
USA. https://birdsoftheworld.org/bow/home.
Claramunt, S., Derryberry, E.P., Cadena, C.D., Cuervo, A.M. Sanín, C.,
and Brumfield, R.T. 2013. Phylogeny and classification of Automolus foliage-gleaners
and allies (Furnariidae). The Condor, 115(2), 375–385, https://doi.org/10.1525/cond.2013.110198
Cory, C. B. and Hellmayr, C. E. 1925. Catalogue of Birds of the
Americas, pt. 4. Field Mus. Nat. Hist. Zool. Ser. 13: 1–390.
Harvey, M.G., et al. 2020. The evolution of a tropical biodiversity
hotspot. Science 370,1343-1348. DOI:10.1126/science.aaz6970
Ridgway, R. 1911. The Birds of North and Middle America, part 5. Bull.
U.S. Nat. Mus. 50: 1–859.
Schultz, E.D., Burney, C.W., Brumfield, R.T., Polo, E.M. Cracraft, J.,
Ribas, C.C. 2017. Systematics and biogeography of the Automolus infuscatus complex (Aves; Furnariidae): Cryptic diversity
reveals western Amazonia as the origin of a transcontinental radiation.
Molecular Phylogenetics and Evolution, 107: 503-515. https://doi.org/10.1016/j.ympev.2016.12.023.
Smith, B., McCormack, J., Cuervo, A. et al. 2014. The drivers of
tropical speciation. Nature 515, 406–409. https://doi.org/10.1038/nature13687.
Oscar Johnson, May 2025
Note from Remsen on English names: If the proposals pass, I see no immediate reason for us to meddle with
English names already chosen by NACC (Chiriqui Foliage-gleaner and Fawn-throated
Foliage-gleaner), but if someone objects, speak up and be prepared to write
separate proposals.
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Remsen:
“A.
[taken from his comments on the original NACC proposal]: YES. Discrimination in playback trials usually
correlates with assortative mating and absence of free gene flow, and so I
interpret these data as showing that these two taxa have differentiated to the
point associated with barriers to gene flow between parapatric/sympatric
species. No one who does playback trials
thinks that this is directly equivalent to mate choice, but rather that it is
an index of potential gene flow if the two were in contact based, empirically,
on many examples that go both ways, i.e. taxa that do not mate assortatively
show strong playback responses, and those that do mate assortatively show low
levels of response. All of this is
discussed in detail in various papers using playback trials. In the imperfect world of assigning taxon
rank to allotaxa, use of playback trials is one of the stronger tools we have,
for better or worse. Much of our current
taxonomy is based on playback trials or degree of divergence in vocal
characters. That professional and
amateur ornithologists worldwide use tape playback to find particular species
provides testimony to the common sense behind interpretation of playback as a
determinant of taxon rank.
“Of
course, in a perfect world we would wait for comparable data between other taxa
in the ochrolaemus complex, but
without a guarantee that such a project is underway, I favor endorsing this
split now. Perhaps this will catalyze
work on other members of the group.
“There
seems to be confusion in terms of what the proposal actually proposes. As in the Recommendation, the allopatric
population of the Chiriquí lowlands is proposed as a separate species, A. exsertus (Chiriqui Foliage-gleaner)
from all other populations of A.
ochrolaemus. As for Pam's question
on whether they are truly allopatric, I assumed that this is the case because
these birds don't get higher than 1400 m, but I have now checked with César
Sánchez (who is doing his dissertation on birds of this region of endemism),
who confirms that they are allopatric as far as is known. The reason hypophaeus remains associated with A. ochrolaemus s.l. is that it is parapatric with other subspecies
treated within A. ochrolaemus; these
form a nearly contiguous set of 3 subspecies that extend from SE Mexico through
Panama across N Colombia. Then, there is
a gap between those populations and the 3 subspecies east of the Andes in
Amazonia, and as noted by Pam, that's where a big change occurs in song. That
the song of hypophaeus may or may not
be closer to nominate ochrolaemus or
other subspecies is a separate issue that should be addressed in future research. As Cadena and Cuervo have shown with Arremon brush-finches in the Andes,
geographic patterns in song variation can show a leapfrog pattern, so
comparisons of these two Costa Rican taxa to the rest of the complex must be
done comprehensively.”
“B. NO,
reluctantly. I can see why Oscar’s recommendation was highly tentative, and I
wonder if WGAC had not already voted on this whether the recommendation would
be different. Using a mtDNA gene tree to
infer phylogeny (from Smith et a. 2014) is just not scientifically valid in
2025, in my opinion. As for the voices,
I can hear a difference, of course, between the few linked recordings. But we’ve debated about with Boesman’s published
analyses were sufficient for taxonomic decisions, so how can we make a taxonomic
decision based, in my opinion, solely (given uncertainty of mtDNA gene tree) on
a couple of recordings without knowing the full variation represented across
the ranges of these taxa? As a matter of
principle, I think we should require better scientific evidence to change the
status quo, even if the status quo is insufficiently grounded in data. This does not mean that I interpret the
current data as indicating BSC conspecificity but rather than the data so far
are inadequate for making a change.
“The two dissenting NACC voters
made these points independently and were not willing to vote for a split based
on a gene tree and unpublished qualitative assessment of a few sonograms. Both emphasized the need to study the contact
zones.”
“The big problem with a NO on this
is that it would leave our broadly ochrolaemus as paraphyletic with
respect to exsertus if the gene trees of Smith et al. (2014) and Schultz
et al. (2017) are species trees; however, I would maintain that whether these
gene trees reflect species trees requires further data, and I would prefer to
maintain a potentially paraphyletic ochrolaemus until those data are
available. I suspect this will be
resolved soon with genomic data from the Harvey lab or elsewhere.”
Comments from Robbins: “
“A. YES. This appears
to be a straightforward decision given playback and genetic data, so a YES for
the split.
“B. YES. Given what we
know, this certainly is more problematic than the recognition of exsertus
as a species because of the lack of vocal data as well as not knowing what
interactions occur in an apparent contact zone between cervinigularis
and ochrolaemus in west-central Panama. Nonetheless, genetic data
(granted only mitochondrial data and we know how misleading that can be) from
two separate studies indicate that cervinigularis appears to be sister
to exsertus. So, if we take those genetic data at face value and
recognize exsertus I think we should go ahead and recognize cervinigularis.
Clearly, additional data may dictate a reevaluation.”
Comments from Lane: “YES to A to align with NACC. I
am further persuaded also to vote YES to B. Vocally, it is obvious that the
Middle American group (cervinigularis/exsertus) is worlds apart
from the South American group (ochrolaemus), and notably, the Chocó
population sounds like the cis-Andean birds and very unlike the Middle American
cervinigularis, not following the usual trend of the Middle American
clade extending to the Chocó. In Panama, I am interested to hear that the song
of pallidigularis is considerably faster than that even from the Chocó.
This is intriguing, but there seems to be a sharpish break between Veraguas and
Panama provinces (the closest recordings I can find between the hypophaeus
and pallidigularis groups, both from Macaulay). I’m hard-pressed to
believe that there is some sort of cline or interbreeding happening in that
zone. Unlike the Troglodytes aedon case, these are suboscines, so
dialect-forming is not likely and the distinctions between the voices are
almost certainly reflected in the phylogenetics, for which we have an mtDNA
tree. It would be nice to have this resolved more explicitly, but I am satisfied
with the combination of datasets we have now to agree to this split.”
Comments from Areta: “When working for AviList/WGAC, I realised
that the 2-way split adopted by eBird, IOC, and NACC seemed problematic, and I
made the following comments on the 13th of December 2022 (synthesized here):
"Wait, if I understand
this correctly, this two-way split would mean two
unacceptable things:
“1) That A.
ochrolaemus would be deeply paraphyletic (because we would be
including the "cervinigularis-amusos-hypophaeus" group with A. ochrolaemus, but it is
sister to exsertus.
See zoom into the image that
Tom posted from Smith et al. 2014:
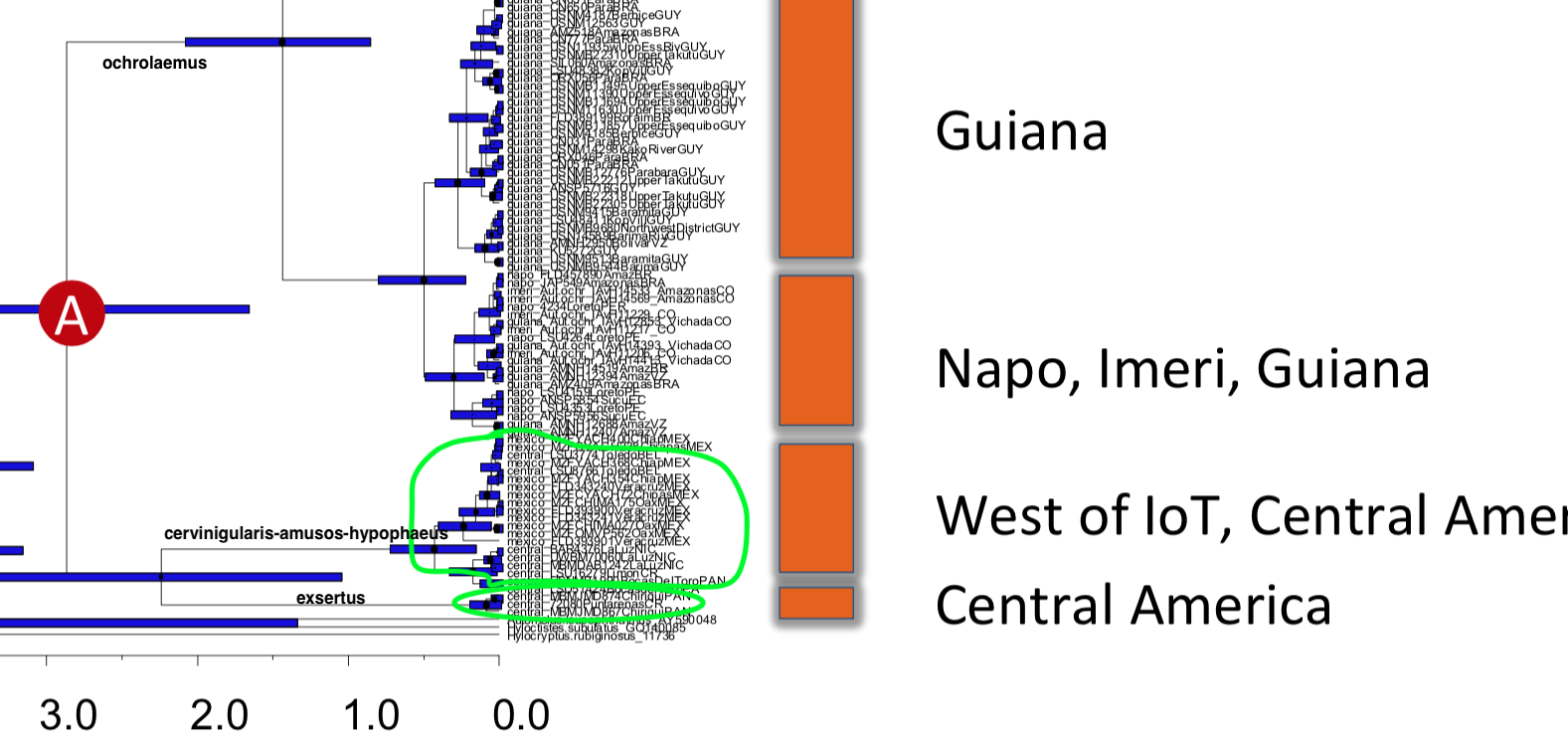
“2) That A.
ochrolaemus would also be vocally very divergent, with two vocal
types clearly identifiable: again, the "cervinigularis-amusos-hypophaeus" vocal type (i.e.,
Mexican) and the nominate ochrolaemus
group (i.e., ochrolaemus-pallidigularis-turdina-auricularis).
--- Take a listen here
“I must say that exsertus and cervinigularis-amusos-hypophaeus
sound more similar among them than to the ochrolaemus
group, exactly as portrayed by the phylogenetic relationships. Listen to exsertus here.
“I cannot access the
comments of the NACC votes (I went through the proposal only).
“I would
therefore vote NO to the two-way split but would be happy to go with a
three-way split, which is consistent with
vocal differentiation and phylogenetic relationships. Either I missed the plot
here, or people have been uncritical to details. I confess that the 3-0
football match today might have had an effect.
“My feeling is that the 2-way split got momentum
because Freeman & Montgomery just did playbacks between two groups."
“I only see a single option here, and it is to adopt
a 3-way split that is consistent with what we know of vocalizations (even
without a formal analysis), and which is also in agreement with available
phylogenetic data:
A-YES
B-YES”
Comments from Stiles: “A. YES to split exsertus;
B. YES to split cervinigularis/hypophaeus from ochrolaemus
(thus 3 species: cervinigularis, exsertus, ochrolaemus).”
Comments
from Zimmer:
“A.
YES. I actually did a lot of
investigation into species-limits in the Automolus ochrolaemus
complex dating back to the 1990s and early 2000s, including borrowing lots of
specimens of all of the named subspecies to measure, photograph and examine
plumage differences. At the same time, I
accumulated an archive of sound recordings from the various taxa to supplement
my own extensive recordings. Alas, this
was one of many such projects that ended up on the back burner as other work
and life priorities took over, and I never got to the stage of writing anything
up, let alone publishing. However, I do
have a lot of pertinent background with this group to bring to the table in
terms of evaluating these Proposals. The
thing that initially stimulated me to investigate species-limits in this
complex, were the vocal differences that I noted between exsertus and hypophaeus
in Costa Rica (and, more broadly, between these Central American birds and all
Buff-throated Foliage-gleaners that I encountered throughout Amazonia and the
Guianas). These differences in song,
although somewhat subtle, were consistent between the Caribbean Slope and
Pacific Slope populations, and that prompted me to conduct playback trials
between the two taxa similar to the ones conducted by Freeman and Montgomery,
2017), nearly 2 decades later. I
conducted the playback trials on hypophaeus from various Caribbean Slope
sites, but primarily from La Selva OTS station and Braulio Carrillo NP. My playback trials on exsertus were
primarily from Las Cruces OTS station (Wilson Botanical Garden) in Costa Rica,
and various sites in Chiriqui Province of western Panama (Pacific Slope). I don’t have access to any of my actual data
at the moment (I’m out of state on vacation as I type this), but I can report
that I had no instances in which individuals of hypophaeus responded to
playback of exsertus, nor any instances in which any exsertus responded
to playback of hypophaeus. There
were rare instances in which an individual Automolus would also exhibit
no appreciable response to playback of same-taxon vocalizations, but in the
vast majority of all trials, foliage-gleaners did respond strongly to playback
of same-taxon songs, while ignoring playback of the allopatric taxon. As such, all of my personal experience with
these two taxa, obtained over 35 years or so of fieldwork in Costa Rica and
Panama, squares with the more systematically conducted and published playback
trials of Freeman & Montgomery (2017), and I strongly agree with their
conclusion and the recent move by NACC to treat exsertus as a separate
species from hypophaeus, and, by, extension, from the remainder of the A.
ochrolaemus-group (more on that in my Part B comments).
“B.
YES. All of my unpublished work on the
complex indicated significant vocal differences between the northern cervinigularis-group
(including hypophaeus) and everything else to the south/east in the
complex, and, as Mark notes, genetic data from two separate studies indicates
that cervinigularis is sister to exsertus. The vocal distinctions
between the cervinigularis-group and the South American ochrolaemus-group
are much greater than the differences between cervinigularis and exsertus,
so much so, that I don’t think we really need a quantitative analysis to rely
upon, given the large number of publicly archived recordings available for
examination. As Dan alludes to, the case
with pallidigularis is particularly interesting. Not only is it vocally closer to the
cis-Andean South American birds than to Middle American cervinigularis,
but it is distinctly whiter-throated than either the northern group of taxa or
the South American group of taxa. Of
particular note, and, again, I am fuzzy on specifics and will have to dig up my
recordings when I’m home, I have a strong recollection of being very excited to
obtain audio recordings of two different vocal types (the normal pallidigularis,
as well as a hypophaeus sounding and looking bird) from the Nusagandi region of San Blas Province sometime prior to
2010. This would establish sympatry
between the two taxa, something I have always meant to follow up on, as part of
the bigger project. At any rate, I think
there’s a decent chance that pallidigularis may prove to be distinct
from the ochrolaemus-group, but I feel confident that it and all of the
South American taxa are different from the cervinigularis-group.
“I
would also agree with Van’s suggestions regarding
English common names if both splits pass (Fawn-throated for cervinigularis;
Ochre-throated for ochrolaemus), and also reserving Pale-throated for pallidigularis,
should that taxon ever be split in the future.”
Comments from Bonaccorso: “A and B: NO. As noted above, it
is logically inconsistent to accept A but reject B, because accepting A alone
would render A. ochrolaemus paraphyletic. The options are therefore to
accept both A and B or reject both.
“The playback experiment offers strong support for the separation
implied by A. However, beyond this result, the remaining arguments rely
primarily on a mitochondrial gene tree, which is not sufficient evidence on its
own. The vocal data is anecdotic and the differences in plumage are so subtle
that it is very difficult to decide based on those lines of evidence.”
“Although I personally suspect that three species are involved, an
integrative taxonomic framework requires multiple independent lines of solid
evidence. At present, the data do not adequately support a three-way split, and
I do not see a compelling reason to rush this decision just to align with WGAC.”
Comments from Naka:
“A: YES. The evidence is compelling, and I am happy to split the
two forms based on song and DNA.
“B: YES, I also agree with the second split, avoiding creating a
monster A. ochrolaemus. I appreciate Nacho and Dan's comments on this
matter.”
Comments from Claramunt: “NO. I agree that playback
experiments do suggest that exsertus may be reproductively isolated from
other members of the complex. But a broader analysis is needed to interpret
that fact in the context of the entire complex. Variation in vocalizations
seems to be compatible with a three-way split, but the evidence so far has not
been properly published. Plumage variation is subtle; there is nothing obvious
there. Genetic data is coming from mtDNA only. In sum, I’m with Elisa: we need
more published evidence in order to properly evaluate species limits in this
complex.”
Additional comments from Robbins: “I also took another look this,
including Santiago's comments. Although I'm now more on the fence about
recognizing these as species, I think comments by Kevin and Dan still have me
in the acceptance camp.”