Proposal (1067) to South
American Classification Committee
Treat Thamnophilus cearensis as a
separate species from Thamnophilus caerulescens
Background:
The
Variable Antshrike (Thamnophilus caerulescens) currently includes eight
subspecies distributed from Peru to eastern Brazil. These taxa have
traditionally been treated as conspecific due to general plumage similarity and
apparent clinal variation among some populations (Zimmer and Isler 2003).
Previous
studies focusing on the western subspecies aspersiventer, paraguayensis,
and dinellii indicated weak reproductive isolation (Brumfield 2005,
Isler et al. 2005, Marcondes et al. 2020). A range-wide genetic analysis,
however, found that the geographically isolated cearensis, from
northeastern Brazil, is deeply divergent from all other taxa
(Bolívar-Leguizamón et al. 2020). Nonetheless, both this and other large-scale
studies (e.g., Marcondes et al. 2020 for plumage) suffered from sparse
geographic sampling, preventing a thorough taxonomic assessment. Moreover,
vocal variation has never been comprehensively studied across the species’
range, despite the widely perceived importance of vocalizations in antbird
taxonomy.
New
Information:
Lima
et al. (2025) conducted an integrative analysis of genome-wide nuclear data,
plumage, morphology, vocalizations, and behavioral responses to song playbacks
across the full range of the Variable Antshrike, using much denser geographic
sampling than any previous study.
They
identified nine phenotypically distinct populations, corresponding to seven of
the eight currently recognized subspecies plus two newly described ones. The following figure illustrates the geographic ranges of
the nine taxa recognized by Lima et al. (2025):
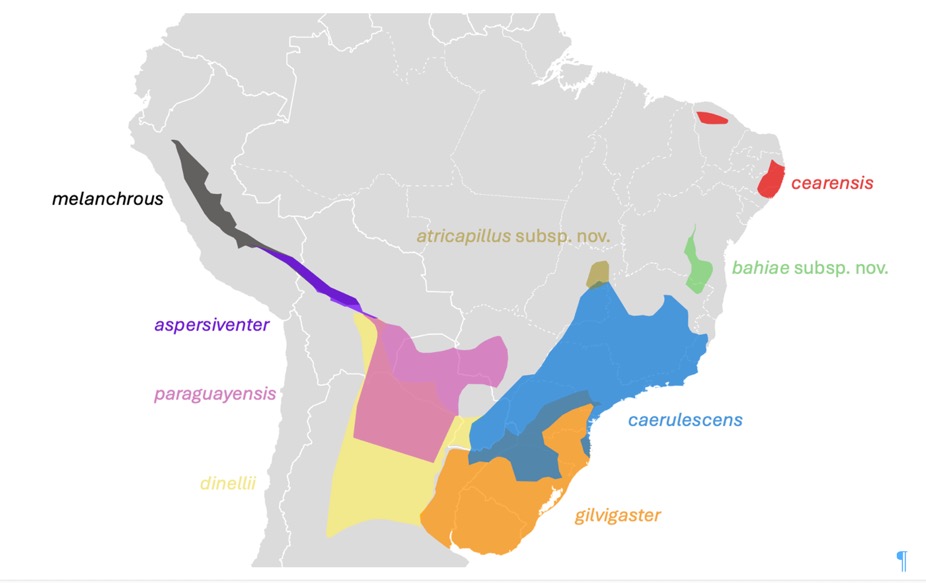
Crucially, Lima et al. (2025) confirmed
that cearensis is noticeably divergent genetically from all other taxa (FST
on the order of 0.24 to 0.38 between cearensis and other taxa) and also
that it has a remarkably distinct song, similar only to that of the
geographically distant melanchrous. In field playback experiments, cearensis
individuals responded to song recordings of its own taxon and melanchrous
while ignoring others.
Recommendation:
The combination of distinct song, discrimination in playback
experiments, and deep genomic divergence indicates that cearensis has
likely evolved substantial reproductive isolation from all other populations of
the T. caerulescens complex. While song-based behavioral isolation
between cearensis and melanchrous appears weak, the two show the
highest pairwise FST values, suggesting that postzygotic
genetic incompatibilities may be strongest between them.
Although direct tests of postzygotic isolation are unavailable,
genetic divergence provides a reasonable proxy. The number of Dobzhansky–Muller
incompatibilities is expected to increase exponentially with genetic distance
between two populations (Orr and Turelli 2001, reviewed in Coyne and Orr 2004).
Consistent with this expectation, Lima et al. (2025) found that parapatric
subspecies with lower genetic differentiation tend to form broader hybrid zones
than those with deeper genetic divergence (see their Fig. 12). Thus, the high
genetic differentiation between cearensis and melanchrous (FST
= 0.38) likely indicates meaningful postzygotic barriers. Inferring
reproductive isolation from this pattern is no more speculative than inferring
behavioral isolation from territorial responses in playback experiments.
Based on this evidence, I recommend a YES vote to treat Thamnophilus
cearensis as a separate species from Thamnophilus caerulescens.
References:
Bolivar-Leguizamón, S.
D., Silveira, L. F., Derryberry, E. P., Brumfield, R. T., & Bravo, G. A.
(2020). Phylogeography of the Variable Antshrike (Thamnophilus caerulescens),
a South American passerine distributed along multiple environmental gradients.
Molecular Phylogenetics and Evolution, 148, 106810.
Brumfield, R. T.
(2005). Mitochondrial variation in Bolivian populations of the Variable
Antshrike (Thamnophilus caerulescens). The Auk, 122(2), 414–432.
Coyne, J.
A., & Orr, H. A. (2004). Speciation. Sinauer Associates.
Isler, M. L., Isler, P.
R., & Brumfield, R. T. (2005). Clinal variation in vocalizations of an
antbird (Thamnophilidae) and implications for defining species limits. The Auk,
122(2), 433–444.
Lima, R. D.,
Bolívar-Leguizamón, S. D., Bocalini, F., Marcondes, R. S., Brumfield, R. T.,
& Silveira, L. F. (2025). Geographic variation, population genetic
structure, and taxonomy of the Variable Antshrike (Thamnophilus caerulescens).
Zoological Journal of the Linnean Society, 205(2), zlaf129.
Marcondes, R. S.,
Stryjewski, K. F., & Brumfield, R. T. (2020). Testing the simple and
complex versions of Gloger’s rule in the Variable Antshrike (Thamnophilus
caerulescens, Thamnophilidae). The Auk, 137(3), ukaa026.
Orr, H. A., &
Turelli, M. (2001). The evolution of postzygotic isolation: accumulating
Dobzhansky‐Muller incompatibilities. Evolution,
55(6), 1085-1094.
Zimmer, K. J. &
Isler, M. L. (2003). Family
Thamnophilidae (Typical Antbirds). In: del J Hoyo, A Elliot, DA Christie (eds),
Handbook of the Birds of the World. Barcelona: Lynx Edicions, 448–681.
Rafael D.
Lima, November 2025
Note from Remsen on English names:
Cory & Hellmayr (1924) called this taxon “Ceará Ant Shrike”. I suggest we simply adopt that sensible name
(as Ceara Antshrike); if anybody has any better ideas, speak out – and do a
separate proposal for the English name.
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments from Remsen: “YES. Vocal differences
with playback experiments make this one an easy decision, in my opinion. That cearensis is the most
isolated taxon, in the complex, geographically, also makes sense.”
Comments from Gustavo Bravo (guest voter): “YES.
Given the vocal, morphological, and genetic evidence, it is appropriate to
recognize Thamnophilus cearensis as a separate species.”
Comments
from Zimmer:
“YES. Way back when the Islers were actively looking into geographic
variation in the T. caerulescens-complex (more focused on the western
array of taxa), and in the lead-up to Mort and I doing the antbird chapter for
HBW, I suggested to Mort that the taxon in the complex to be most focused on
was cearensis, which was not only the most isolated taxon, but which, to
my ears, was vocally distinct from all other Brazilian taxa in the complex, and
which showed the most divergence in female plumage as well. I even did some informal, one-way playback
trials in the Serra de Baturité, employing only songs of nominate caerulescens
and gilvigaster, and found, like Lima et al (2025), that cearensis
did not respond to playback of either caerulescens or gilvigaster
vocalizations, but did respond aggressively to playback of pre-recorded
vocalizations of other cearensis.
I never did reciprocal playback trials of cearensis songs to
either caerulescens or gilvigaster populations, nor did I do any
trials involving any of the western taxa in the caerulescens-complex. So, nothing as rigorous or comprehensive in
scope as what Lima et al (2025) have published, but this is just to say that
all of my own fieldwork with this complex is in line with their conclusions. Assuming this passes, I would also be
strongly in favor of Van’s suggested English name of “Ceara Antshrike” for T.
cearensis.”
Comments
from Naka:
“YES. I agree with the new treatment, which was long overdue.”
Comments
from Stiles:
“YES - evidence from genetics, phenotype, and distribution justify this split.
Comments
from Robbins:
“YES, given the new data.”
Comments
from Lane:
“YES. It looks like all datasets agree with splitting these two groups.”
Comments from Areta: “A
fascinating system and a great study by Lima et al. 2025. I am however mostly
not satisfied with the taxonomic decisions of the work. It is of course
impossible to tackle all the fronts of this widespread species at once, and
while some results are clear, others are not (not to me at least).
“cearensis
as a species: plumage differences vs the closest taxon (newly
described bahiae)
are very minor and limited to females. The only "Isler-Whitney"
difference between the geographic closer cearensis
and bahiae is on
note shape, which falls short of the 3 such characters that set the standard
level of differentiation in order to be considered different species in the
family by the same Isler-Whitney team. The playback experiments (Table S3) are
suggestive, but overall inconclusive: nearly 50% of the caerulescens tests did not
discriminate, cearensis
could not discriminate against melanchrous,
and the number of playback stimuli indicates rampant problems of
pseudoreplication by lack of adequate number of stimuli. More rigorous playback
experiments are needed. Of course, one can choose to ignore methodological
problems and take an "overall" message, but I don´t think that the
overall message is that clear either. The Fst between the newly described bahiae and cearensis is on par with
those between other populations (especially comparisons involving the other
geographically extreme taxon melanchrous)
which we are not considering as a species-level candidate. The deepest
differentiation between melanchrous
and cearensis (the
two more distant and disconnected taxa) is quite high, but they are bridged by
intermediate populations. Finally, do the phylogenetic trees recover bahiae and cearensis as reciprocally
monophyletic? It seems from Figure 11, that cearensis
is embedded within bahiae.
I might be alone in this world, but I
vote NO to the split of cearensis.
I am comfortable with considering it as an allopatric population of the
widespread caerulescens,
being similar vocally and in plumage to its closest (yet allopatric) neighbour,
bahiae. It is
clear that different populations show different degrees of isolation and
hybridization, and cearensis
is the most extreme one but not yet commensurate to species level
differentiation by Thamnophilus
standards.
“The two
new subspecies described have diagnostic females, but not
diagnostic males, which cannot be distinguished from neighbouring populations.
Given the impressive number of specimens analyzed, I would have expected heavy
doses of photographs instead of drawings. Not even the holotypes were pictured!
I am not enchanted by the descriptions.
“atricapillus
(new taxon #1): This taxon seems hard to justify, despite its apparent bold and
easily diagnosable female aspect, in which "They are closest in appearance
to females of caerulescens and gilvigaster, from which they
differ by having a black crown" (Lima et al. 2025: 16). First of all, this
subspecies would have a minute range (or even no proper range), and furthermore
Lima et al. 2025 (16) stated that "we documented plumage intergradation
between caerulescens and new taxon #1 in central Brazil (Fig. 7).
Although the contact zone appears narrow (∼31 km), phenotypic intergradation
spans the entire range of new taxon #1 (Fig. 7). ". So, if phenotypic
intergradation spans the entire range, then I don´t see how we can delimit a
range for this taxon whose full range is an intergradation zone between itself
(an intergrade) and another taxon. Confusing. In looking at the specimen table
(Table S1) I find 20 females "f_new1" phenotype individuals and 52
males "m_new1_or_caerulescens" phenotype individuals. Although the
diagnosis indicates the taxa from which males are diagnosable, it does not
mention those from which they are not. If the phenotypic intergradation spans
the entire range of the new taxon, then it seems like this taxon does not have
an independent existence and is therefore questionable and fuzzy. I vote NO to the recognition of atricapillus, mostly
because of how confusingly the situation was described.
“bahiae
(new taxon #2): the females are extremely similar to those of cearensis, and males are
indistinguishable from the latter. Females of bahiae "are closest
in appearance to females of cearensis, from which they differ by being
more brownish and less olivaceous on the belly, breast, back, and wing
coverts" (Lima et al 2025: 16). I can accept this taxon, on account of the
difference of females from the neighbouring caerulescens.”
Additional
comments from Remsen: “After reading Nacho’s much more thorough
comments on this, I feel bad for not having seen those points myself, and
change my vote to NO. Also, this has no
bearing on my view of the quality of the paper itself – in general, the quality
of the paper is independent from the taxonomic conclusions from it.”
Comments
from Claramunt: “Reluctant YES. I truly commend Rafael and co-authors
for such a thorough study with great sample sizes and an integrative approach.
But there is no clear picture emerging from the data and analyses regarding the
species status of cearensis. The genetic analyses presented in this
study are not totally convincing. It is mostly based on methods borrowed from
numerical taxonomy (PCA, k-means clustering). I would like to see more
model-based phylogenetic and population genetic analyses of this complex to
determine potential relationships and levels of gene flow. PCA on discrete data
such as SNPs always tends to exaggerate differences among group. The
phylogenetic network in Figure 11 is the closest to a phylogenetic analysis and
the long branch leading to cearensis suggests that it might be the most
basal branch, but without outgroup, Nacho’s interpretation of cearensis deriving
from bahiae is a plausible alternative. Bolívar-Leguizamón et al.
produced a mtDNA tree in which cearensis is indeed the most distantly
related lineage in the complex.
“According to the taxonomic summary, females of cearensis“
differ diagnostically in plumage from all other taxa” but I could not find the
evidence for this statement. The cited Fig. 1 shows only illustrations of
idealized individuals in which females of cearensis and bahiae look
indistinguishable. The diagnosis of bahiae indicates that they are
diagnostically browner, less olivaceous, than cearensis, but the term
“diagnostic” is used very liberally in this study with most “diagnosable taxa”
having wide “transition” zones and polymorphisms that are incompatible with
strict diagnostic status.
“However, all things considered, the phylogenetic data
plus partial diagnosability in plumage and song mildly suggest that cearensis may
be better treated as a full species.”
Additional
comments from Robbins: “After reading Nacho's comments and
revisiting the paper (especially the taxonomic conclusions), I agree with his
conclusion, that at best, it would be premature to recognize cearensis as a
species. Thus, I change my vote to NO.”
Additional
comments from Lane: “Thanks to Nacho for the extensive response, and I must
confess I didn’t really parse the case well in my first round vote as I was a
bit pressed for time, but I did listen to some recordings and was not
particularly impressed with the distinctiveness of cearensis compared to
my own experience with various populations (especially in the Andes) of T.
caerulescens. So, I feel less silly about not seeing cearensis as so
distinctive, and I will change my vote to NO with regard to the proposed split.”