Proposal (1072) to South
American Classification Committee
Recognize new species of Hylophilus
INTRODUCTION
A geographically isolated taxon of Hylophilus
has been known for quite some time from Beni, Bolivia (Olrog 1960), ca. 2000 km
away from the nearest population of the morphologically similar H.
amaurocephalus and H. poicilotis. Different authors have chosen to
treat the new population as either H. amaurocephalus or H. poicilotis,
without settling on a definitive treatment (Herzog et al. 2014), as H.
amaurocephalus (Ridgely & Tudor 1989) or as a putative new taxon (Lane
2014, van Els et al. 2024).
Based on Van Els et al. (2026), we recommend
species rank for H. moxensis based on: 1. Relatively deep genetic
divergence between H. moxensis and H. amaurocephalus/H. poicilotis (the
latter two currently recognized species based, among others, on sympatry, see
Willis 1991), 2. Diagnostic morphological differences on the head, particularly
the plain auriculars compared to H. amaurocephalus/H. poicilotis, 3.
Geographic isolation over ~2000 km precluding gene flow, and 4. Habitat
differences between H. moxensis and H. amaurocephalus/H. poicilotis.
We propose the English name Beni Greenlet, which is
appropriate in view of its known geographic range, which is confined to the
department of Beni, Bolivia.
RATIONALE
Firstly, we found a well-supported relatively deep
divergence (Fig. 1, 6.6±2.5–11.0 mya) between both H. amaurocephalus and
H. poicilotis (which diverged 3.7±1.6–5.4 mya from each other, albeit
with low nodal support) and the Beni birds. We used the same four (ND2, ACOI,
MUSK, SPIN1) genes as those employed in a broader phylogeny of Vireonidae by
Slager et al. (2014) and we included all fully-sampled species of Hylophilus
from Slager et al. as well as Cyclarhis gujanensis as an
outgroup.
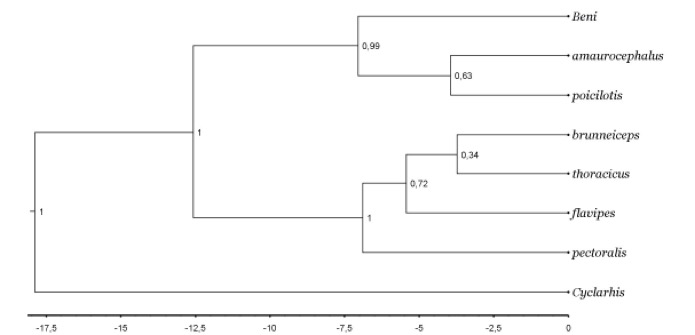
Fig.
1. Bayesian species tree of genus Hylophilus including samples from
Slager et al. (2014) and novel data, based on ND2 (1041 bp), ACO1 (967
bp), MUSK (448 bp) and SPIN (616 bp). Notice the well-supported divergence
between H. moxensis and H. amaurocephalus/poicilotis.
We investigated three existing specimens and collected
an additional two specimens to investigate morphological differences with H.
amaurocephalus and H. poicilotis. This is a relatively modest sample
size, because permitting restrictions precluded us from collecting more
specimens. Morphometric measurements of the few available specimens largely
overlapped with those of H. amaurocephalus and H. poicilotis,
although the largest culmen and tarsal measurements of H. moxensis
exceeded those of H. poicilotis.

Fig.
2. Typical head patterns in H. poicilotis, moxensis and amaurocephalus
and Discriminant Correspondence Analysis of discretized scores indicating
morphological groupings.
To increase sample size for analytical purposes and to
be able to build upon the previous methodology by Raposo et al. (1998),
we used a set of discrete morphological characters related to the head (some of
the same characters scored by Raposo et al. (1998) and found to be
distinguishing H. amaurocephalus and H. poicilotis, plus
additional characters) and scored these from a large number of photos
available, as well as specimens in case of the Beni, Bolivia birds. The three
taxa of Hylophilus in the study are very similar in coloration over most
of the body; and while we acknowledge there may be differences in e.g. the
intensity of yellow on the underparts between the three, we focused on
characters related to the head, eyes and bills of the three, which can be
relatively easily discretized (and are therefore not so dependent on light
conditions as would be continuously distributed characters such as color
saturation or value). Scored character states were discretized with minimal
ambiguity in state assignment; most characters were therefore coded using just
three states to avoid over-categorization (auricular coloration using four).
Consistent with the genetic distinctness of the three lineages, they are also
morphologically diagnosable.
We found (just like Raposo et al. 1998) that H.
amaurocephalus has a dark gray grizzling on the auriculars, which is more
extensive and generally fused to a dark auricular patch in H. poicilotis.
H. moxensis is characterized by having uniform auriculars without any
dark grizzling (except for a postocular spot or line, Fig. 2). Like northern H.
amaurocephalus and H. poicilotis, H. moxensis always has dark
eyes. On average, H. moxensis also has more dark on the commissure and
lores than H. amaurocephalus or H. poicilotis. According to all
characters taken together in multivariate analysis, the three groups are
fully diagnosable as distinct entities. We did not find any apparent geographic
variation in morphology of H. moxensis.
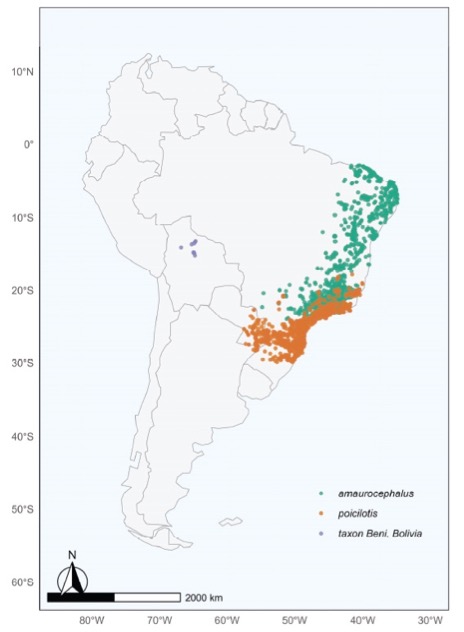
Figure
3. Verified records of H. amaurocephalus (green), H. poicilotis
(orange) and H. moxensis (purple).
We provide additional qualitative information on
vocalizations, habitat and distribution. Although potentially diagnostic,
vocalizations of Hylophilus show a large amount of geographic and
individual variation, as well as variation related to level of agitation, so
even with a large number of recordings, species-specific diagnostic
vocalizations are hard to establish. See also Raposo et al. 1998 for the
impressive amount of individual and geographic variation in songs and calls of
the two sister species of H. moxensis, just as that found in other
Corvides (see Rheindt et al. 2022). We found that H. moxensis has
harmonics in its calls, just as do female H. amaurocephalus, but unlike
H. poicilotis. In spite of their large and more geographically varied
sample of vocalizations of both of H. moxensis’ sister species, this
was apparently the only diagnostic feature found by Raposo et al. (1998).
Songs of H. moxensis may have upward or downward inflections at the end,
whereas Raposo et al. (1998) found that H. poicilotis generally
have upward and H. amaurocephalus have downward inflections, with
exceptions.
We know of no populations of Hylophilus in the H.
amaurocephalus/poicilotis complex in the areas in between Beni, Bolivia and
the Brazilian ranges of H. amaurocephalus/poicilotis and the assertion
that the latter two may have migratory populations (e.g. Herzog 2014) that may
wander westward seems unfounded. The Beni birds are restricted to gallery
forest and scrub in the seasonally inundated savannas of Beni, Bolivia, and
have yet to be found in other semi-arid habitats in surrounding regions (e.g.
Chiquitania, Pantanal, Chaco, Fig. 3) and so are more restricted in their
habitat choice than either of the two Brazilian species, which may be found in
semi-arid scrub to humid forest and edge.
RECOMMENDATIONS
A1.
Recognize H. moxensis as a full species. A 'NO' vote on proposal A1
would require votes on the following alternative treatments: (A2) treat moxensis
as a subspecies of H. poicilotis, or (A3) treat moxensis as a
subspecies of H. amaurocephalus.
B.
Adopt Beni Greenlet as the English name of this taxon.
A 'YES' vote is recommended for proposals A1 and
B.
Literature
Herzog S.K., Terrill R.S., Jahn A.E., Remsen J.V.,
Maillard O.Z., García-Solíz V.H., MacLeod R., Maccormick A., Vidoz J.Q., 2016. Birds
of Bolivia: field guide. 1–491. – Asociación Armonía, Santa Cruz, Bolivia.
Lane D.F., 2014. New and noteworthy records of birds
in Bolivia. – Cotinga, 36: 56–67.
Olrog C.C., 1963. Notas sobre aves bolivianas. – Acta
Lilloana, 19: 407–478.
Raposo M.A., Parrini R. & Napoli M., 1998.
Taxonomia, morfometria e bioacústica do grupo específico Hylophilus
poicilotis / H. amaurocephalus (Aves, Vireonidae). – Ararajuba,
6 (2): 87–109.
Rheindt F.E., Wu M.Y., Movin N., & Jønsson, K.A.,
2022. Cryptic species-level diversity in Dark-throated Oriole Oriolus
xanthonotus. –Bulletin of the British Ornithologists’ Club, 142(2),
254-267.
Slager D.L., Battey C.J., Bryson Jr R.W., Voelker G.
& Klicka J., 2014. A multilocus phylogeny of a major New World avian
radiation: the Vireonidae. – Molecular Phylogenetics and Evolution, 80:
95–104.
van Els P., Wijpkema T., Wijpkema J.T. &
Montenegro-Avila M., 2024. Changes in the status and distribution of savanna
birds of Beni and Santa Cruz, Bolivia. – Cotinga, 46: 2–13.
van Els P., Montenegro-Avila M., Avilos, N.A.,
Wijpkema T. & Wijpkema J.T., 2026. A new species of greenlet from Bolivia
in the Hylophilus poicilotis/amaurocephalus group (Vireonidae). – Avian
Systematics 3: 17–37.
Willis, E. O. 1991. Sibling species of greenlets
(Vireonidae) in southern Brazil. – The Wilson Bulletin, 559-567.
Paul van Els, March 2026
Vote
tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Stiles: “YES; genetic, phenotypic, vocal and distributional data
all suggest its distinctiveness. And YES to Beni Greenlet - given the very
distinctive distribution, to the south of the ranges of other greenlets, this
effectively singles out this species without the need to search through the
genus to derive a distinctive plumage-based name.”
Comments
from Robbins: “YES because of genetic data and habitat differences. The
difference in plumage is so slight to be almost meaningless.”
Comments
from Lane:
“YES. This was a taxon that had been on my radar for
almost two decades, in which time I did playback experiments, made sound
recordings, and tried to encourage LSU expeditioners to collect specimens in an
effort to advance the study. It is distinctive in plumage, voice, habitat, and
structure when compared to the H. poicilotis/amaurocephalus
group. I also approve of the chosen names by the authors.”
Comments from Areta:
“A.1.YES. The vocal, plumage, and genetic data
support the recognition of H. moxensis
as a new species. The genetic divergence seems very deep for such modest
phenotypic differences.”
“A.2. NO
“A.3. NO
“B. YES, Beni
Greenlet is accurate and helps single this taxon out. Given its late entrance
into the game, the geographic modifier seems reasonable.”
Comments from
Rasmussen: “B. YES. I too vote for Beni. It
seems to be endemic to the department, which does not currently have any bird
species named for it as far as I can see. Hard to see any downside to this
name!”
Comments
from Donsker: “B. I vote YES for Beni Greenlet. In my opinion it's without
question the most appropriate name for this restricted range species.”
Comments from
Claramunt: “YES. Subtle but fully
diagnostic plumage differences plus the phylogenetic evidence demonstrate that
the Beni population is a species-level taxon. Geographic isolation should not
play a role as evidence; neither ‘habitat differences,’ as it is very difficult
to infer intrinsic habitat preferences for birds that live in regions far
apart.”
Comments from
Remsen:
“A. YES. Species rank is clearly
the best interpretation of the genetic and vocal information. I echo Santiago’s comments on the lack of
relevance of geographic isolation and habitat preferences in this case.”
“B. YES. Good choice, and “Beni”
makes sense in English (instead of “El Beni”).