Proposal (1075) to South
American Classification Committee
Split Tufted Flycatcher
Mitrephanes phaeocercus into two or three species and modify English
names
Note: This proposal
is here repurposed for SACC, having been considered by NACC.
Summary
Although vocal differences have formed
the basis for many recent changes in species limits within Tyrannidae, the
striking vocal differentiation within the Mitrephanes phaeocercus
complex has not yet been formally considered as a basis for revision of the
species limits, although it has been known for some time (see for example Dyer
and Howell 2023). Tufted Flycatchers in northern Middle America have position
calls and dawn songs that differ strikingly from those of birds from Costa Rica
south, and this division aligns well with plumage differences and is supported
by evidence of a strong genetic break between Honduras and eastern Panama. The
differences in position note are less striking between Costa Rica/Panama (M.
p. aurantiiventris) and e. Panama/Colombia/Ecuador (M. p. berlepschi),
but the differences in primary song, dawn song, and plumage strongly suggest
that those should also be recognized at the species level. The only other
described taxon in Mitrephanes (there is a putatively undescribed taxon
on Cerro Tacarcuna), Olive Flycatcher Mitrephanes olivaceus, is already
recognized as specifically distinct. Acceptance of this proposal would require
change in English names of the daughter taxa of phaeocercus, and we
recommend a minor change to that of M. olivaceus as well.
This proposal does not give statistical
measurements of the songs and calls, in part because the differences are so
striking: the position notes are so dissimilar between phaeocercus and aurantiiventris
that it is challenging to find relevant measurements. The Macaulay Library (ML)
provides ample audio recordings to support the vocal differences, and
supporting links are provided below.
Background
The
genus Mitrephanes is thought to be
closely related to Contopus (pewees),
which includes multiple species that are nearly identical in appearance and
differ primarily in vocalizations.
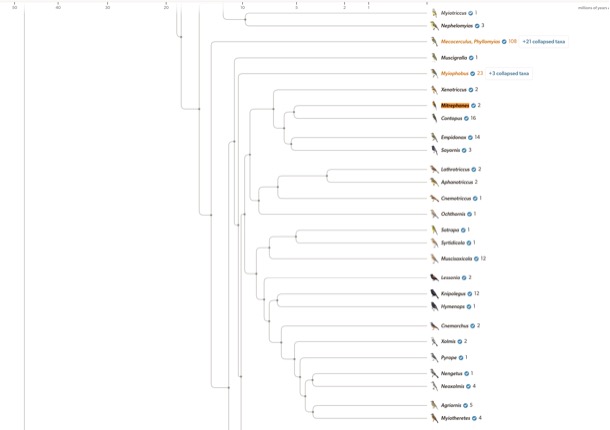
Fig.
1.
The Birds of the World Phylogeny Explorer shows the position of Mitrephanes
as sister to Contopus; see https://birdsoftheworld.org/bow/phylogeny#tuffly; Miller et al.
(2025).
Webster
(1968) summarized the taxonomic history within Mitrephanes as follows:
“Ridgway (1907) recognized five species, although he was able to
examine only two. Hellmayr (1927) lumped the entire genus in a single species,
as did Zimmer (1930; 1938a) in the most thorough revision thus far. Griscom
(1932) restated the specificity of M. berlepschi, and Sutton and
Burleigh (1940), while they did not discuss the South American forms,
distinguished M. aurantiiventris (of southern Central America) as a
species separate from M. phaeocercus. It should be noted that Zimmer had
only 76 specimens altogether and Sutton and Burleigh only 38 from north of the
Isthmus of Tehuantepec.”
The
AOS-SACC has followed the recommendation of Webster (1968), which was to
recognize two species within Mitrephanes. AOS-NACC considered the genus
monotypic until 2018, when it finally voted to conform with the AOS-SACC
interpretation by recognizing M.
olivaceus (Olive Flycatcher), which is extralimital to the AOS-NACC area.
That 2018 proposal (Chesser 2018; https://americanornithology.org/wp-content/uploads/2020/02/2018-A.pdf) was
straightforward and explicitly recommended following Webster (1968) and the
AOS-SACC interpretation.
Thus,
as of 2018, both AOS-NACC and AOS-SACC follow the recommendations on species
limits from Webster (1968) wherein he recommended “A moderate course between
the extreme points of view, which recognize four species or one, seems best to
express the phylogeny of the genus. I would recognize two species--M.
phaeocercus and M. olivaceus.”
Webster
(1968) recognized all the below taxa in Mitrephanes; eBird/Clements and
AviList treat at least four subspecies in synonymy, as below; taxa recognized
by eBird/Clements (Clements et al. 2025) and AviList (AviList Core Team 2025)
are listed in bold type. Ranges are modified from those in eBird/Clements.
o
M. p. tenuirostris Brewster,
1888: mountains of western Mexico (southeastern Sonora and
southwestern Chihuahua to western Jalisco)
o M. p. phaeocercus (Sclater,
1859): mountains of eastern and central Mexico (southwestern
Tamaulipas, Zacatecas, and central Jalisco to Oaxaca and Chiapas) southward to
northeastern Nicaragua
§ M. p. burleighi Phillips, 1966: central Jalisco to w. Oaxaca
§ M. p. phaeocercus (Sclater,
1859): mountains of eastern and central Mexico (southwestern Tamaulipas and
Zacatecas southward to e. Oaxaca
§ M. p. nicaraguae Miller & Griscom,
1925: northern Chiapas to northeastern Nicaragua
o M. p. aurantiiventris (Lawrence,
1865): highlands of Costa Rica and Panama (eastward to Serranía del
Darién*)
§ M. p. aurantiiventris (Lawrence,
1865): highlands of Costa Rica and Panama (eastward to e. Chiriquí and
Ngäbe-Buglé provinces)
§ M. p. vividus Griscom, 1927: central
Panama (highlands of e. Coclé and w. Panamá; affinities of birds in Veraguas is
unclear); treated as synonym of aurantiiventris in eBird/Clements and
AviList.
o M. p. berlepschi Hartert,
1902: far eastern Panama to northwestern Colombia and northwestern
Ecuador
§ M. p. eminulus Nelson, 1912: eastern
Panama (Darién, including Cerro Pirre and Tacarcuna ranges) and presumably far
nw. Colombia
§ M. p. berlepschi Hartert,
1902: northwestern Colombia and northwestern Ecuador
o
M. olivaceus Berlepsch & Sztolcman, 1894: east
slope of Andes of northeastern Peru (Piura) to northwestern Bolivia
*“Serranía
del Darién” is evidently incorrectly included in the range of aurantiiventris
in Clements et al. (2025 and previous) and will be modified to “(eastward to
Altos del Maria area in eastern Coclé and western Panamá provinces)” in the
2026 Clements update.
Thus,
eBird/Clements and other authors treat burleighi and nicaraguae
as synonyms of phaeocercus, vividus as a synonym of aurantiiventris,
and eminulus as a synonym of berlepschi. eBird/Clements has long
recognized these subspecies groups within the species:
·
Mitrephanes phaeocercus phaeocercus/tenuirostris Tufted Flycatcher
(Mexican)
· Mitrephanes phaeocercus
aurantiiventris Tufted Flycatcher (Costa Rican)
· Mitrephanes phaeocercus
berlepschi
Tufted Flycatcher (South American)
Below,
we refer to these three groups as phaeocercus Group, aurantiiventris,
and berlepschi; when differences between phaeocercus and tenuirostris
are relevant, the specific subspecies name is used. Note however that Webster
(1968) treated nicaraguae as a distinctive plumage class separate from phaeocercus/tenuirostris.
Genetics
We
are not aware of a robust genetic study with complete taxon sampling of the key
groups within Mitrephanes, but Harvey et al. (2020) sampled M.
olivaceus (San Martín, Peru; La Paz, Bolivia), M. p. phaeocercus
(Honduras), and M. p. berlepschi (Darién, Panama). Their time-calibrated
tree (Table 1 in their supplementary material) estimated the divergence times
as follows: Mitrephanes from Contopus (~5.75 mya), M.
phaeocercus/berlepschi clade from M. olivaceus (3 mya), M. p.
phaeocercus from M. [p.] berlepschi (2 mya). The divergence time
between berlepschi and phaeocercus is thus estimated to be older
than or in a similar range to several other well-established species pairs in
their study, including:
· Empidonax traillii and E. alnorum
(~1.5 mya)
· E. atriceps and E. fulvifrons
(1.75 mya)
· E. affinis and E. oberholseri
(1.5 mya)
· E. flavescens and E. difficilis
(~2 mya)
· Contopus pertinax, C. lugubris,
and C. fumigatus (~1.75 mya)
· C. cinereus and C. nigrescens
(1.5 mya)
· Sayornis phoebe and S. nigricans
(~1.75 mya)
The
fact that aurantiiventris was not sampled leaves a significant question
about how divergent it may be from berlepschi and the phaeocercus
Group, but it is clear that M. phaeocercus and M. berlepschi
should be treated within different species and more than meet a genetic
yardstick for related species in Tyrannidae.
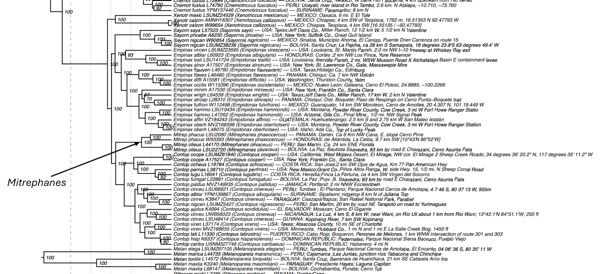
Fig. 2. Table 1 from Harvey et al. (2020) shows a
divergence time of ~2 million years ago between Mitrephanes in Honduras
(M. p. phaeocercus) and Darién, Panama (M. p. berlepschi), which
is consistent with treating these as different species. See also https://tree.opentreeoflife.org/curator/study/view/ot_2015
Habitat
Significant
differences in habitat between at least two of these taxa. Webster (1968)
describes habitat differences as follows:
“My own field
experience with Mitrephanes has been confined to México. There M.
phaeocercus inhabits the more humid parts of the pine-oak forest and cloud
(subtropical) forest. In winter it lives in tropical deciduous forest, tropical
evergreen forest, and tropical thorn forest as well as in lower-elevation pine
forest and cloud forest; it deserts higher elevation pine forest. All that I
encountered in pine or pine-oak forests were foraging out from exposed perches
at the rims of barrancas or mountain ridges. In cloud forest they were foraging
out over openings or low trees and returning to perches 20 to 100 feet above
the ground, in the manner of a pewee (Contopus).
“Skutch (1960)
states that from Guatemala to western Panama the Tufted Flycatcher is a
permanent resident of subtropical forest and of temperate pine-oak forest. He
describes the behavior briefly, including evidence of a close relationship with
the pewees. All localities in central and eastern Panamá reported in the
literature and on specimen labels (M. p. viridus and M. p. eminulus)
are from elevations of 2,000 feet or more, apparently in the subtropical zone.
But the few localities in which M. p. berlepschi has been found, in
western Colombia and northwestern Ecuador, are near sea level in the humid
tropical zone! On the other hand, M. olivaceus in Peru and Bolivia
occurs in upper humid tropical and subtropical forests at elevations from 4,000
to 8,000 feet (Zimmer, 1938a).”
Although
the habitat differences between the phaeocercus Group and aurantiiventris
in part follow the changeover in montane forest type, it does seem notable that
berlepschi is more of a lowland taxon than aurantiiventris.
Morphometrics
Webster
(1968) provided an in-depth phenotypic analysis, working from 728 museum
specimens from all named taxa and from all regions throughout the range of Mitrephanes
(except Ecuador) to quantify plumage and morphology. He (Webster 1968) provided
detailed information on the measurements of the various populations in Mitrephanes.
In general, these follow Bergmann’s Rule, with measurements increasing with
distance from the equator. The full tables for measurements are provided in his
paper.
Webster
noted these specific observations that he considered significant:
· wing length: smooth
cline from northeastern Mexico to western Panama, but sharp break between
Colombia and Peru with a nonoverlap gap of 4mm (over twice the standard
deviation), and lesser breaks (with slight overlap) between Colombia and
eastern Panama and between central and western Panama.
· tail length: sharpest
break between Honduras-El Salvador and Guatemala
· bill width: two
smallest (shortest-winged) populations, vividus and berlepschi,
have the broadest bills
Thus,
the morphometrics given by Webster (1968) do not align particularly well with
the recognized subspecies groups.
Plumage
For
a group of birds in which plumage differences can be extremely subtle (e.g.,
Eastern vs. Western wood-pewees, various taxa of Tropical Pewee, Alder vs.
Willow flycatchers, etc.), the plumage differences among the four main groups
of Mitrephanes are striking. Underparts coloration is especially
strikingly different between the groups, but differences also exist in
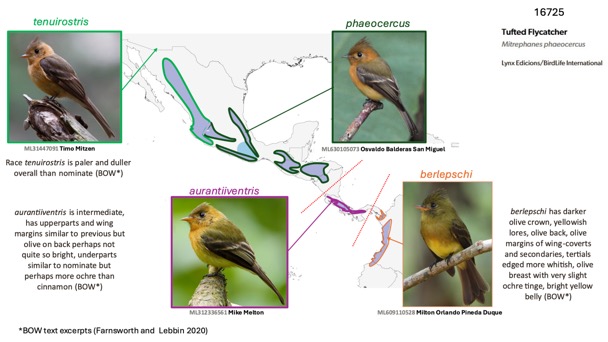
upperparts coloration. The composites below show the differences well (see also
Appendix A for photo galleries from different regions):

Fig. 3. Clockwise from
top left: tenuirostris, Quinn Diaz, Nayarit, MX, ML 626188382; aurantiiventris,
Gary Rosenberg, Panamá, PA, ML610105559; berlepschi, Eric VanderWerf,
ML610105559, Panamá, PA; olivaceus, Itamar Donitza, ML646534080, Cusco,
PE.

Fig. 4. Clockwise from top
left: tenuirostris, Dylan Osterhaus, ML632797034, New Mexico, US; aurantiiventris,
Dubi Shapiro, ML281494981, Chiriquí, PA; berlepschi, Guillermo NAGY /
ARAMACAO TOURS, ML625759985, Chocó, CO; olivaceus, Eric Atwell, ML
612973767, Junín, PE.
Webster
(1968) assessed fresh-plumaged birds (Aug-Feb) and attempted to quantify the
differences, and in doing so he identified five classes (which include two
classes in the northern group from Mexico south to Nicaragua). [Note that while eBird/Clements and IOC
previously recognized nicaraguae, these were synonymized with phaeocercus
around 2005, and that treatment has carried over to AviList v2025. However,
the analysis by Webster (1968) would seem to support recognizing nicaraguae
at least; see below.]
Webster
(1968) noted five classes for underparts coloration, with 100% separation: (1)
pale tawny--tenuirostris, burleighi, and phaeocercus; (2)
dark tawny--nicaraguae; (3) buffy yellow--aurantiiventris and vividus;
(4) yellow--eminulus and berlepschi; (5) lime--olivaceus.
Below
is a summary of the differences demonstrated by Webster (1968):
phaeocercus Group
· underparts pale tawny (tenuirostris,
burleighi, and phaeocercus) to dark tawny (nicaraguae)
· moderate and gradual
transition between anterior underparts (throat and breast) and posterior
(belly, crissum, and undertail coverts)
· Upperparts varying from pale to dark olive and pale to dark brownish
olive
· Juvenal plumage: Ventrally a smooth cline from pale buffy
brown in Sonora darkening southeastwardly to deep tawny buffy brown to brownish
tawny in Chiapas and Nicaragua, while dorsally a smooth cline ran from pale
sepia tipped with pale tawny in Sonora, darker southeastward to dark sepia
tipped with tawny in Chiapas, Guatemala, Nicaragua, and Honduras
· moderate clinal
variation detected, with general increasing darkness to the south
aurantiiventris
· underparts buffy
yellow—aurantiiventris and vividus
· prominent and sharp
transition between anterior underparts (throat and breast) and posterior
(belly, crissum, and undertail coverts)
· upperparts approximately dark, yellow lime-olive
· Juvenal plumage underparts slightly paler and distinctly
yellowish buffy brown, and dorsally the upperparts were paler (than phaeocercus
Group) and had more yellowish tawny tips
· no geographic variation
detected in color
berlepschi
· underparts yellow—eminulus
and berlepschi
· prominent and sharp
transition between anterior underparts (throat and breast) and posterior
(belly, crissum, and undertail coverts)
· in underparts
coloration, two specimens from Cerro Tacarcuna, eastern Panama, are warmer,
more orangish on both areas than two specimens from southern Colombia. In this
class the anterior underparts are medium, brownish Buffy Yellow; the posterior
are pale Yellow.
However,
note that seven worn old (March-June, 1912 and 1928) specimens from Cana,
eastern Panama, show distinct variation; they are colder, less buffy than those
from Cerro Tacarcuna, although paler, less greenish anteriorly and less purely
yellow posteriorly than specimens from Colombia, either northern or southern.
· upperparts varying from medium to dark olivaceous
Yellow-Lime
olivaceus
· underparts lime
· moderate and gradual
transition between anterior underparts (throat and breast) and posterior
(belly, crissum, and undertail coverts)
· Juvenal plumage tawny buffy brown below, paler and buffier
(than aurantiiventris), buffy-brown tipped tawny above
· little geographic
variation, including between Peru and Bolivia
Webster
(1986) furthermore stated:
“The gap between groups 2 [aurantiiventris] and 3 [berlepschi]
is prominent, with the eastern Panamá birds distinctly greener than those from
western and central Panamá.
I see no geographic variation within the 39 specimens from Costa Rica
and western Panamá, and the gap from class (2) [nicaraguae] is
considerable.
Two specimens from Cerro Tacarcuna, eastern Panamá, are warmer, more
orangish on both areas than two specimens from southern Colombia. In this class
the anterior underparts are medium, brownish Buffy Yellow; the posterior are
pale Yellow. Seven worn old (March–June, 1912 and 1928) specimens* from Cana,
eastern Panamá, show a distinct variance; they are colder, less buffy than
those from Cerro Tacarcuna, although paler, less greenish anteriorly and less
purely yellow posteriorly than specimens from Colombia, either northern* or
southern.”
See
Appendix B regarding the possibility of an undescribed taxon on Cerro
Tacarcuna.
Regarding
the upperparts, Webster (1986) stated:
“A prominent north to south cline of increasing greenness runs from
northwestern Mexico to Peru. In Mexico north of Tehuantepec the trend is from
the Pacific slope to the Atlantic slope, but in Central America no Pacific
versus Atlantic slope differences are apparent.
A two-way cline of darkness decreases from maximum in Chiapas northwest
and southeast to minimum, or pallor, in northwestern Mexico and Peru. This
seems to follow Gloger's Rule north from Chiapas, but to proceed almost
inversely to, Gloger's Rule from Chiapas southward.”
We
recommend reading the full text of Webster (1968: https://digitalcommons.usf.edu/cgi/viewcontent.cgi?article=18739&context=auk).
Webster
(1968) assessed juvenal plumage and found marked differences between the phaeocercus
Group, aurantiiventris, and olivaceus:
“Color of juvenal plumage.—A series of 43 specimens from all subspecies
except vividus and berlepschi showed prominent geographical
variation. Ventrally a smooth cline from pale Buffy Brown in Sonora darkened
southeastwardly to deep tawny Buffy Brown to brownish Tawny in Chiapas and
Nicaragua; in Costa Rica and eastern Panamá the underparts were a little paler
again and distinctly yellowish Buffy Brown; in Peru tawny Buffy Brown.
Dorsally, a smooth cline ran from pale Sepia tipped with pale Tawny in Sonora,
darker southeastward to dark Sepia tipped with Tawny in Chiapas, Guatemala,
Nicaragua, and Honduras; in Costa Rica and eastern Panama the upperparts were a
little paler again and had more yellowish Tawny tippings. In Peru the
upperparts were still paler and buffier, Bully Brown tipped with pale Tawny.”
Webster's
(1968) review focused on measurements, appearance, and habitat, and from that
he recommended a two-species treatment. However, his only defense for the
treatment as two species is his section on SPECIFIC LIMITS on page 288–289. The
entirety of his argument is reproduced below:
“All the forms of Mitrephanes are allopatric. When Zimmer (1930)
described the juvenal plumage of Peruvian M. olivaceus as more similar
to that of the Central American forms than is the adult plumage, he argued that
this showed their essential conspecificity. As shown in the section on
variation below, present evidence shows distinct discontinuities in variation.
The most abrupt break occurs in Ecuador, between the coastal forest of western
Colombia and northwestern Ecuador on the one hand and the eastern slope of the
Andes in Peru on the other. Lesser breaks occur in Central America--one between
Nicaragua and Costa Rica, and another between central and extreme eastern
Panama.
“A moderate course between the extreme points of view,
which recognize four species or one, seems best to express the phylogeny of the
genus. I would recognize two species--M. phaeocercus and M. olivaceus:
|
|
phaeocercus |
olivaceus |
|
Range |
Northern Mexico to
northwestern Ecuador |
Eastern Peru and Bolivia |
|
Habitat |
Temperate pine and
pine-oak forest, subtropical forest, and humid tropical forest |
Subtropical forest and
upper humid tropical forest |
|
Size |
Larger to smaller |
Larger than adjacent
forms |
|
Color |
Brown to yellowish green |
Green—contrast marked
with phaeocercus ventrally, but moderate dorsally |
If
an intermediate population exists, it may probably be found in the central
Andes of Colombia.”
It
strikes us that this is an exceptionally weak argument for both AOS-NACC and
AOS-SACC to use as the basis for their species-level taxonomy. Although we
agree that the separation of olivaceus as a distinct species is the
correct course, we stress that Webster (1968) acknowledged the possible
alternate arrangement and made no argument for or against it.
An alternate hypothesis, using Webster’s
own phenotypic data, might look like this:
|
|
phaeocercus |
aurantiiventris |
berlepschi |
olivaceus |
|
Range |
Northern Mexico to
Nicaragua |
Costa Rica to central
Panama |
Eastern Panama to n. Ecuador |
Eastern Peru and Bolivia |
|
Habitat |
Temperate pine and
pine-oak forest and subtropical forest |
Humid tropical forest in
highlands |
Humid tropical forest in
lowlands |
Subtropical forest and
upper humid tropical forest |
|
Size |
Larger |
Moderate-sized |
Small |
Larger than adjacent
forms |
|
Color of underparts |
Pale tawny to dark tawny;
moderate breast-belly contrast |
Buffy yellow; strong
breast-belly contrast |
Yellow; strong breast-belly
contrast |
Lime; moderate
breast-belly contrast |
|
Color of upperparts |
Pale to dark olive or
brownish-olive |
dark, yellow lime-olive |
varying from medium to
dark olivaceous yellow-lime |
varying from medium to
dark olivaceous yellow-lime |
An
equal, or stronger case, could be made that the populations with the traits
above merit recognition as four species, and that when the strong vocal break
between phaeocercus Group and aurantiiventris, and the moderate
vocal break between aurantiiventris and berlepschi, are taken into account, one must arrive at a three- or
four-species treatment.
The
below graphic shows typical phenotypes on the combined range for the taxa
currently lumped as Tufted Flycatcher (M. phaeocercus sensu lato). The
breaks (red dotted lines) between northern Nicaragua and Costa Rica and between
central and eastern Panama are breaks that define numerous other species limits
of highland taxa:
Vocalizations
Comparing
homologous vocalizations is of course of major importance with flycatchers,
with their innate vocalizations. Below, we identify six main classes of
vocalizations, but focus especially on position note, primary song, and dawn
song, which seem to show species-level differences among each of the four
groups. See also Appendix A for galleries of sound recordings from different
regions.
Position
note:
repeated, commonly given series either while perched or immediately after
landing. These tend to be repeated in rapid succession from 3x to 12x or more
Primary
song:
short, high-pitched phrases of single, well-spaced notes; these are given
primarily in the breeding season
Alternate
song:
combining elements of the position note and the song, these tend to be phrased
a pip-pip-pip-seeeu or something similar
Dawn
song:
given pre-dawn or just after dawn, these are longer and complex series but have
well-established phrase patterns
Contact
calls:
various pip notes; occasionally burry interaction calls as well; these
do not seem particularly distinctive and are similar between taxa
Other
calls:
occasionally burry interaction calls and other less consistent vocalizations
may be given as well; these do not seem particularly distinctive and are
similar between taxa
The
vocalizations of the four main groups of Tufted Flycatchers are substantially
different and more than exceed the vocal differences between many flycatcher
species. We did not detect significant differences across the ranges of any of
the taxa, although a quantitative analysis may shed more light on this aspect.
Most
striking is the difference in position notes: those of phaeocercus are
utterly unlike the position notes of any of the other taxa. The position notes
of aurantiiventris and berlepschi are similar, and while those of
olivaceus are similar in pattern, they have a very different note shape,
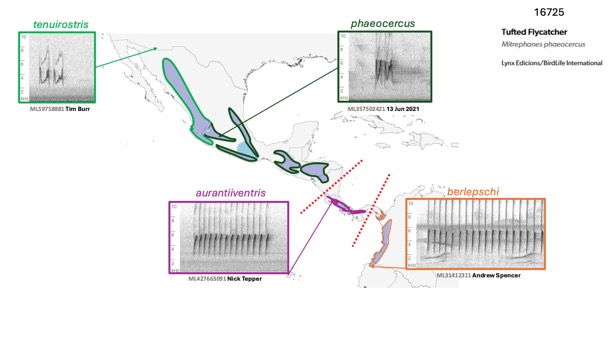
being descending vs. ascending. Figure 5 summarizes the differences.
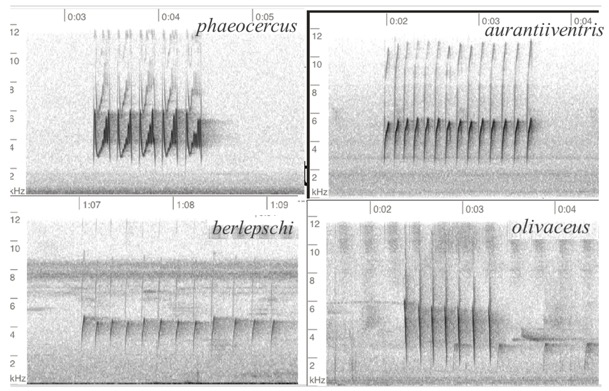
Fig. 5. Position
calls. Clockwise from top left: phaeocercus, Cd. de Mexico, MX, Miguel
Aguilar @birdnomad, ML650288423; aurantiiventris, Alajuela, CR, Nick
Tepper, ML427663091; olivaceus, Amazonas, PE, Guilherme Melo,
ML608680193; berlepschi, Valle de Cauca, CO, Paula Caycedo, ML264163.
Below, graphic showing typical position calls of taxa in M. phaeocercus s.l.
The
primary songs (below) are all a similar short, high-pitched note and reportedly
are given primarily in the breeding season. However, the shape of the notes
differs substantially between the taxa, with more similarity between phaeocercus
and aurantiiventris (despite the massive differences in position calls),
while berlepschi and olivaceus differ substantially both from
each other and from the two more northerly taxa.
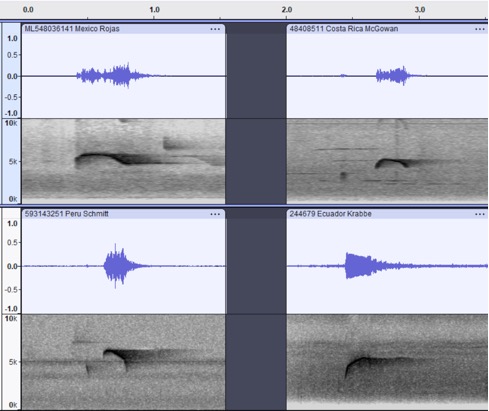
Fig. 6. Primary song.
Clockwise from top left: phaeocercus, Querétaro, MX, Alvaro Rojas Qro.
Birding Tours, ML548036141; aurantiiventris, Cartago, CR, Jay
McGowan, ML48408511; berlepschi, Esmeraldas, EC, Niels Krabbe, ML244679;
olivaceus, San Martín, PE, Fabrice Schmitt, ML593143251.
Vocalizations
that we refer to as alternate songs combine elements of the position call and
the primary song. It seems not be given frequently in the phaeocercus
Group, or potentially the preceding notes are reduced to a single pip
with an element of the position call (https://macaulaylibrary.org/asset/109246). The other
three taxa all have multiple examples of the alternate song, with multiple pip
notes preceding a high-pitched element that is analogous to the primary song.

Fig. 7. Alternate
song. From the top: aurantiiventris, Puntarenas, CR, Thomas Brooks,
ML590274961; berlepschi, Esmeraldas, EC, Mark L. Robbins, ML63198; olivaceus,
Amazonas, PE, Nick Athanas, ML288261481.
The
dawn songs are audio-recorded much less frequently, but the several examples
for each taxon show differences in pattern, speed, and quality of delivery that
suggest that each taxon is a species. The final notes for each phrase seem to
be an especially important element of the dawn song in these taxa and do not
have much similarity between the phaeocercus Group, aurantiiventris,
and berlepschi.
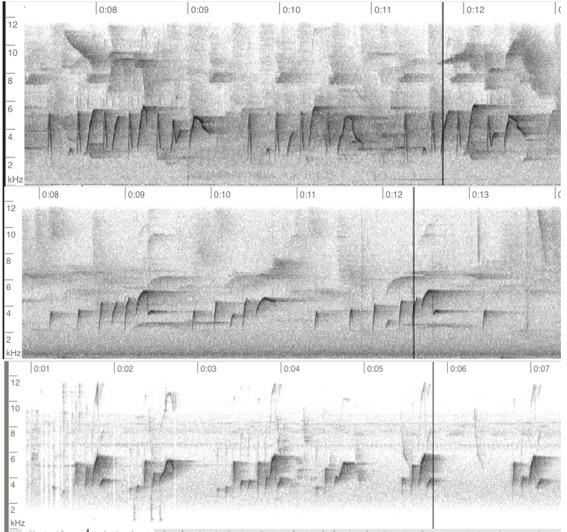
Fig. 8. Dawn song.
From the top: phaeocercus, Hidalgo, MX, Andrew Spencer,
ML277665301; aurantiiventris,
Puntarenas, CR, David L. Ross, Jr., ML184565; berlepschi, Valle de
Cauca, CO, Daniel Uribe-Restrepo BIRDING TOURS COLOMBIA, ML90360501. Below, a
graphic mapping a different selection of dawn-type songs for the taxa of M.
phaeocercus s.l.
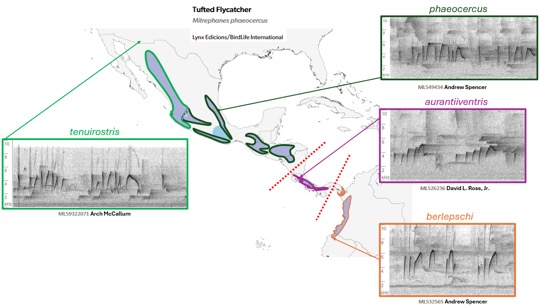
We
found no obvious examples of dawn song of M. olivaceus on Macaulay
Library or xeno-canto; the one example of a complex song was more like a
complex version of the alternate song and was quite unlike the other three
taxa: https://xeno-canto.org/20777
The
below table summarizes the differences in vocalization. The alphanumeric codes
in parentheses refer to the vocal analyses in Appendix C and identify the
vocalization type and taxon.
|
|
context |
phaeocercus grp. |
aurantiiventris |
berlepschi |
olivaceus |
|
position notes |
Loud series given perched
or just after landing; elements repeated 3x or up to 10x or more; a very
common vocalization in the genus |
strident, burry
tchwee-tchwee- tchwee (A1) |
loud and rapid, repeated
series pui-pui-pui-pui… (A2) |
loud and rapid, repeated
series pee-pee-pee-pee… (A3) |
loud and rapid, repeated
series tip-tip-tip-tip… (A4) |
|
primary song |
High-pitched single note
that varies in pitch; given in breeding season and often repeated every few
seconds; not unlike like song given by many Contopus |
high, thin rising and
falling pseeeu; dome-shaped
spectrogram with peak near front of note (B1) |
high, thin rising and
falling seer or peew; symmetrical, dome-shaped spectrogram or
sometimes with peak at the front (B2) |
wheep (B3); rising note that levels off at the end |
sieeuu (B4); fast peak and then long descending note |
|
alternate song |
Series that combines
short elements from the position call with the primary song or a modified
primary song |
rarely given, perhaps
absent; pip-weet (C1) |
pip-weer (C2) |
tsu-tsu-tseét (C3) |
tip-tip-tip-tip-seeert, with strong descending note (C4) |
|
dawn song |
Complex series covering a
wide frequency range, given primarily at dawn |
variable phrases of 4-7
notes, pit-whit-ooh-whee-ohh… pit-whit-ooh-whit-wheet… with last
element alternating between whee-ohh (L-shaped note) and weeet (scythe-shaped
note) |
variable phrases of 4-7
notes, pit-whit-ooh-whee-weeot… pit-whit-ooh-whit-wheet… with last
element alternating between double-humped wheeot (double-humped
descending note) and weeet (scythe-shaped note) |
variable phrases of 4-7
notes, pit-whit-ooh-whee-weeot… pit-whit-ooh-whit-wheet… with last
element alternating between double-humped wheeot (double-humped
descending note) and weeet (scythe-shaped note) |
no examples found |
|
contact calls |
Short single “pip” notes,
recalling Alder or Hammond’s Flycatcher; given frequently and similar in each
taxon to a lone element of the position call |
pip, peaking around 5 kHZ (E) |
pip, peaking around 4 kHZ (E) |
pip, peaking around 4 kHZ (E) |
tip, peaking around 5 kHZ (E) |
Field
guide descriptions are as follows:
Howell
and Webb (1995) - phaeocercus Group: A burry, bright,
rolled tchwee-tchwee or turree-turree, less often a single tchwee
or longer series, a sharp, usually fairly quiet pic or beek
suggesting Hammond’s Flycatcher, and a bright, slightly emphatic to penetrating
seeu or pseeu, repeated every few s and given mainly in breeding
season.
Howell
et al. (2014) - phaeocercus Group: Bright, slightly burry
whistled tch’wee-tch’wee or tureee-turee call is given
year-round, as well as quiet pik, suggesting call of Hammond’s
Flycatcher.
Stiles
and Skutch (1989): aurantiiventris Group
A
high-pitched, rapid series of liquid, rather confiding notes: weet weet weet…or
pui-pui-pui-pui-pi-pi , often upon returning to perch; a thin seer
or peew or, especially in breeding season, peew-peew or peep-wit
wit; quaint, prolonged dawn song a very rapid series of high, thin
notes: bip-bip-bip-dididi-up-bip-bip-bibibiseer.
Wetmore
(1972):
aurantiiventris - Eisenmann describes their calls as a piping pip-pip-pip-pip
varying in the number of notes and occasionally uttered at a faster pace.” He
remarks that the notes suggest those of the much larger Dark Pewee but are
higher in pitch.
Angehr
and Dean (2010): Call a rapid series of high chirping pip notes
[presumably aurantiiventris Group, no distinction made between western
and eastern birds]
Ridgely
and Gwynne (1989) – berlepschi Group: Frequently gives a
piping whistle, pee-pee-pee (up to eight notes).
Ridgely
and Greenfield (2001) – berlepschi Group: Call an often-heard
fast series of pee or pik notes. Song a repeated fast phrase of
high, thin notes, e.g., tsu-tsu-tseét, with softer notes sometimes
interspersed between phrases (P. Coopmans).
Hilty
and Brown (1986) – berlepschi Group: Frequently repeats a
spirited bubbly ser. of 4-7 pee or pik notes as flicks tail; call
reminiscent of that of Greater Pewee but faster, weaker, and not as sharp.
Effect
on NACC area:
If successful, this proposal would result in the recognition of either one or
two additional species for the region.
Recommendation
Mitrephanes
olivaceus
is already recognized as a distinct species, and we recommend no change to that
classification; it is included above to allow for yardstick comparisons to the
other taxa.
Within
the current Tufted Flycatcher M. phaeocercus there are three vocal
groups that match the described subspecies, show well-defined plumage breaks
(as described by Webster 1968), and match well-known biogeographic regions
(Mexico to Nicaragua, Costa Rica/western Panama, and eastern Panama to northwest
Ecuador, west of the Andes).
The differences in vocalizations (especially
contact call and dawn song) are especially striking between Nicaragua and Costa
Rica but are much more similar between central Panama and e. Panama/Colombia. A
conservative option would be to recognize M.
phaeocercus and M.
aurantiiventris, with berlepschi included as a subspecies of M.
aurantiiventris.
However, it is also clear that aurantiiventris and berlepschi show fairly strong plumage differences between central and eastern
Panama and also have substantial differences in primary song and dawn song
(although reduced differences in position calls relative to other taxa). For
many other taxa of flycatchers, vocal differences of this magnitude would be
sufficient to warrant species status, and we think that given the correlated
differences in plumage, and the different elevational preferences and habitat
of berlepschi, these should be treated as separate species as well.
Whether
to split phaeocercus (with tenuirostris) from the other taxa is
primarily a NACC issue, but both this split and whether to further split berlepschi
from aurantiiventris affect SACC inasmuch as the name berlepschi
is junior to both other names. The options for SACC thus are:
OPTION
A:
Recognize three species:
· Mitrephanes phaeocercus from Mexico to n. Nicaragua
· Mitrephanes aurantiiventris from w. Costa Rica to
central Panama
· Mitrephanes berlepschi from e. Panama to nw. Ecuador, west of
the Andes
OPTION
B:
Recognize two species:
· Mitrephanes phaeocercus from Mexico to n. Nicaragua, including M.
p. tenuirostris from nw. Mexico as a subspecies
· Mitrephanes aurantiiventris, from w. Costa Rica to n. Ecuador,
including M. p. berlepschi from
e. Panama to nw. Ecuador as a subspecies
English names: If this proposal passes, a separate proposal may be needed to address
English names. Dyer
and Howell (2023) have used Mexican Tufted Flycatcher for M. phaeocercus
sensu stricto and Costa Rican Tufted Flycatcher for M. aurantiiventris.
Cinnamon and Tawny Tufted-Flycatcher could also be used for M. phaeocercus
s.s. and Isthmian or Yellow-bellied Tufted-Flycatcher could be used for M.
aurantiiventris, and Choco Tufted Flycatcher for M. berlepschi (if
split). Habitat-based names would also be possible, with Pinewoods or
Pine-forest Tufted-Flycatcher for M. phaeocercus and Cloud-forest
Tufted-Flycatcher (with Choco Tufted-Flycatcher for M. berlepschi).
Should the latter two not be split, many of these English name options would
not be appropriate, and perhaps Northern and Southern Tufted-Flycatcher should
be considered.
However, we strongly recommend adopting as a group
name either Tufted Flycatcher or Tufted-Flycatcher, which would align with
AviList v2025. IOC (Gill et
al. 2025) and AviList have used Northern Tufted Flycatcher for Mitrephanes
phaeocercus s.l. and Olive Tufted Flycatcher for M. olivaceus,
while AOS-NACC, AOS-SACC, and BirdLife International (e.g., del Hoyo and Collar
2016) have used Tufted Flycatcher for M. phaeocercus and Olive
Flycatcher for Mitrephanes olivaceus. If this proposal passes and Mitrephanes
expands from a two-species genus to a genus with three or four species, then it
would be a good time to consider whether it would be better to follow IOC and
AviList and use “Tufted-Flycatcher” for all members of the genus.
Just
in the New World, there are 105 species across 31 genera that bear the name of
"flycatcher". For the lay birder, it certainly can be hard to keep
track of which English name goes with which genus without knowing the bird very
well. Only the two Royal Flycatchers have a name that helps tie them together
and distinguish from the other tyrannids. If the Old World is considered, yet
another 151 species across 22 additional genera are involved, although Crested
Flycatchers, Blue Flycatchers, and Jungle Flycatchers do help to orient birders
toward assemblages of congeneric, or at least similar, species. While adopting
Tufted Flycatcher would not solve the core problem of too many unrelated birds
with the name of “flycatcher”, it would help birders to orient correctly on Mitrephanes
and to transfer familiarity more readily from one species to another.
Tufted
Flycatcher is also a highly appropriate name that would be a shame to lose with
this revision. While a lot of tyrannids have elongated crown feathers that can
be raised as a crest, there may not be any others with such long and pointed
crests as Mitrephanes. For a notoriously difficult-to-identify family, Mitrephanes
is wonderfully straightforward to identify precisely because of their sharply
pointed crests that are usually fully raised. We strongly feel that maintaining
the most distinctive feature of the genus within the name of all its members
follows best practices for naming.
Especially
relevant to SACC, Olive Flycatcher is an extremely ambiguous and
non-distinctive name for M. olivaceus—how many tyrannids could be
described as olive? Renaming it to Olive Tufted Flycatcher or Olive
Tufted-Flycatcher would be a significant improvement, as it is the only Mitrephanes
that is aptly described as olive. When NACC first recognized Olive Flycatcher
as distinct in 2018 (Proposal set 2018-A) six voters
opted not to use a compound name Tufted-Flycatcher, most citing the fact that
SACC was not doing so. Three voters felt it was important to not use something
as ambiguous as Olive Flycatcher and to follow IOC in using Olive
Tufted-Flycatcher. Assuming passage of this proposal, there would be 3-4
species in this genus, it feels even more important to use the compound name to
tie all Mitrephanes together as Tufted-Flycatchers. Therefore, we
recommend the use of Tufted Flycatcher or Tufted-Flycatcher for all species of Mitrephanes.
In
hopes of a speedy resolution, we recommend the following names for SACC-region
species (YES or NO):
Choco
Tufted-Flycatcher Mitrephanes berlepschi
Olive
Tufted-Flycatcher Mitrephanes olivaceus
Literature
Cited
Angehr,
G. R. and R. Dean (2010); The Birds of Panama: A Field Guide. Comstock, Ithaca,
NY, USA.
AviList
Core Team (2025). AviList: The Global Avian Checklist, v2025. https://doi.org/10.2173/avilist.v2025
Chesser, R. T.,
K. J. Burns, C. Cicero, J. L. Dunn, A. W. Kratter, I. J. Lovette, P. C.
Rasmussen, J. V. Remsen, Jr., D. F. Stotz, B. M. Winger, & K. Winker
(2018). Fifty-ninth Supplement to the American Ornithological Society’s Check-list of North American Birds. Auk 135: 798–813.
Clements,
J. F., P. C. Rasmussen, T. S. Schulenberg, M. J. Iliff, J. A. Gerbracht, D.
Lepage, A. Spencer, S. M. Billerman, B. L. Sullivan, M. Smith, and C. L. Wood.
2025. The eBird/Clements checklist of Birds of the World: v2025. Downloaded
from https://www.birds.cornell.edu/clementschecklist/download/
del
Hoyo, J., and N. J. Collar (2016). HBW and BirdLife International Illustrated
Checklist of the Birds of the World. Volume 2: Passerines. Lynx Edicions,
Barcelona, Spain.
Dyer, D. and S. N. G. Howell (2023). Birds of Costa Rica. Princeton Field Guides.
Princeton University Press, Princeton, NJ, USA.
Farnsworth, A. and D. J. Lebbin (2020). Tufted
Flycatcher (Mitrephanes phaeocercus), version 1.0. In Birds of the
World (J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana,
Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.tuffly.01
Gill, F., D. Donsker, & P. C. Rasmussen. Eds. 2025. IOC World Bird List (v 15.1).
http://www.worldbirdnames.org/
Harvey, M. G. et al. (2020). The evolution of a tropical
biodiversity hotspot. Science 370: 1343–1348.
Hilty,
S. L. and W. L. Brown (1986). A Guide to the Birds of Colombia. Princeton
University Press, Princeton, NJ, USA.
Howell, S. N.
G. and S. Webb (1995). A Guide to the Birds of Mexico and Northern Central
America. Oxford University Press, Oxford, UK.
Howell, S. N.
G., I. Lewington, and W. Russell (2014). Rare Birds of North America. Princeton
University Press, Princeton, NJ, USA.
Miller, E. T., E. J. McTavish, J. A. Gerbracht, M.
Schloss, M. J. Iliff, D. Lepage, P. C. Rasmussen, and B. L. Sullivan (2025).
The Phylogeny of the Birds of the World. Tree v1.6/Taxonomy v2025. Cornell Lab
of Ornithology, Ithaca, NY, USA.
Ridgely,
R. S. and J. A. Gwynne (1989). A Guide to the Birds of Panama. With Costa Rica,
Nicaragua, and Honduras. Princeton University Press, Princeton, NJ, USA.
Ridgely,
R. S. and P. J. Greenfield (2001). The Birds of Ecuador. Cornell University Press, Ithaca, NY, USA.
Stiles, F. G. and A. Skutch (1989). A Guide to the
Birds of Costa Rica. Cornell University Press, Ithaca, NY, USA.
Webster, J. D. (1968). A revision of the Tufted
Flycatchers of the Genus Mitrephanes. The Auk 85(2): 9. Available at: https://digitalcommons.usf.edu/auk/vol85/iss2/9
Wetmore, A. (1972). The Birds of the
Republic of Panama - Part 3. Passeriformes: Dendrocolaptidae (Woodcreepers) to
Oxyruncidae (Sharpbills). Smithsonian Institution Press, Washington, D.C., USA.
Appendix A: Media
galleries
Audio
Galleries:
Mexico:
https://tinyurl.com/w6e5scm6
Northern
Central America + Chiapas: https://tinyurl.com/4rx9evau
Costa
Rica and Panama (except Darién): https://tinyurl.com/4s8mk2ur
Darién
and Colombia: https://tinyurl.com/msaz8wrs
Photo
Galleries:
Tufted
Flycatcher (South American): https://tinyurl.com/mrhvthuz
Tufted
Flycatcher (South American), Cerro Pirro area, Panama: https://tinyurl.com/34ve4jx2
Tufted
Flycatcher, Tacarcuna: https://tinyurl.com/47hpzzzx
Tufted
Flycatcher, El Valle: https://tinyurl.com/rdvetx5z, https://tinyurl.com/yc3hecj5, and https://tinyurl.com/mr5ks26d
Costa
Rica: https://tinyurl.com/5xvjdrva
Appendix
B. Undescribed taxon at Cerro Tacarcuna?
As
noted by Webster (1986), comparison of photos from Cerro Pirre with those from
Cerro Tacarcuna in Darién show plumage differences that suggest there is an
undescribed taxon on Cerro Tacarcuna.



In
the above image, the three images from the Cerro Tacarcuna range (outlined in
orange) suggest that there are significant morphological differences between
the birds in the Cerro Tacarcuna area (e.g., https://macaulaylibrary.org/asset/42900761) and those
from Cerro Pirre (e.g., https://macaulaylibrary.org/asset/144776101), which match
images of berlepschi well from nearby areas in Colombia. The birds from
the Tacarcuna region appear to have rich cinnamon-rufous breasts, which extends
to the nape, and a whitish belly, while typical berlepschi (and the
Cerro Pirre) completely lack cinnamon or rufous tones, and instead show bright
lime green bird with a yellow breast lightly washed with orange and pale yellow
belly. Our suspicion is that there exists an undescribed taxon in the Tacarcuna
area. The general coloration, with cinnamon breast contrasting with a paler
belly best matches aurantiiventris (which has a yellowish belly) and we
recommend provisionally treating this taxon within that species until the
Tacarcuna birds can be described and their species limits assessed with
vocalizations, a more detailed study of morphology, and ideally, genetics.
Appendix
C. Detailed comparison of vocalizations
The
below analysis of vocalizations (by MJI) attempts to use the context of
vocalizations to cluster them in analogous groupings. Each vocalizations
equates roughly with the songs vs. calls of related taxa (e.g., Contopus)
which have high pitched simple songs, more complex dawn songs that incorporate
elements of the song, and calls that often involve single or repeated pip
calls. The set of vocalizations termed the position note match the pip-pip-pip
calls given by Greater Pewee but seem to be given even more frequently.
When
vocalizations differ significantly between taxa, I have split those as
vocalization 1, 2, 3. Thus, in the example below all examples of Vocalization A
are believed to be analogous contact calls. These calls are among the most
commonly given, heard year-round, and all are often given immediately after
birds return to perches. With the exception of A1 (phaeocercus Group),
all have obvious audio similarities.
VOCALIZATION A1 –
Position calls
Taxa: Mexican phaeocercus
Group
Context: position call,
given commonly, especially right after landing
Description: burry, breeep
given singly, (often) doubled, less commonly delivered 3x, 4x or 5x.
Howell
and Webb (1995) describe this as a “burry, bright, rolled tchwee-tchwee or turree-turree,
less often a single tchwee or a longer series” while Howell et al.
(2014) describe it as a “bright, slightly burry whistled tch’wee-tch’wee
or tureee-turee call is given year-round”
Discussion: This
vocalization is entirely unique within the group and greatly exceeds the vocal
differences between many other flycatchers considered different species. The
difference here is on par with the vocal differentiation between the bik
and twitter calls of Couch’s and Tropical Kingbirds, for example. The notes are
burry, often doubled and rarely given in longer series, and completely unlike
the analogous calls (repeated pip series) for all other taxa in the
genus. It seems hard to argue that this vocalization from a member of
Tyrannidae could be conspecific with aurantiiventris or berlepschi.
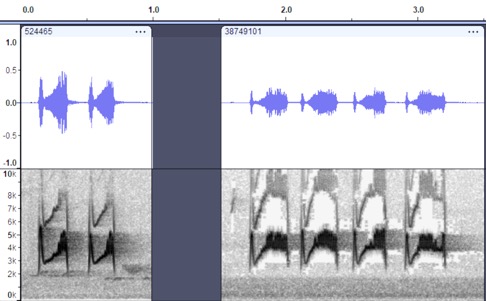
Nayarit,
MX https://macaulaylibrary.org/asset/524465 Nayarit, MX (delivery 3x or 4x) https://macaulaylibrary.org/asset/38749101 or even 5x
https://macaulaylibrary.org/asset/130035001
https://macaulaylibrary.org/asset/64233221
Recordings from the far south of the range of phaeocercus
(in Nicaragua) are clearly still of this call type:
https://macaulaylibrary.org/asset/523408
VOCALIZATION A2 –
Position calls
Taxa: aurantiiventris
Context: position call,
given commonly, especially right after landing
Description: repeated pip
notes, often trailing off a bit at the end (more spacing between notes,
descending somewhat in pitch). Described by Wetmore (1972) for aurantiiventris
as “Eisenmann describes their calls as a piping pip-pip-pip-pip varying
in the number of notes and occasionally uttered at a faster pace.” He remarks
that the notes suggest those of the much larger Dark Pewee but are higher in
pitch.” Angehr and Dean (2010) “Call a rapid series of high chirping pip
notes” [no distinction made between western and eastern birds] and Ridgely and
Gwynne (1989) say “frequently gives a piping whistle, pee-pee-pee… (up
to eight notes).” Stiles and Skutch (1989) give the best description of
vocalizations “a high-pitched, rapid series of liquid, rather confiding notes: weet
weet weet…or pui-pui-pui-pui-pi-pi, often upon returning to perch.
Hilty and Brown (1986) state: “Frequently repeats a spirited bubbly ser.
of 4-7 pee or pik notes as flicks tail; call reminiscent of that
of Greater Pewee but faster, weaker, and not as sharp.”
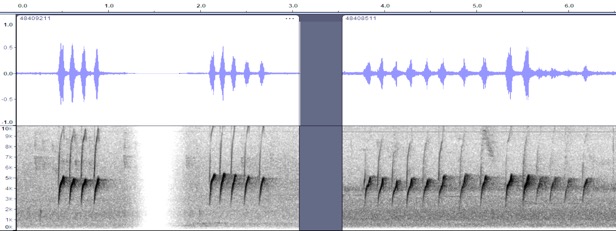
repeated
4x or 5x, https://macaulaylibrary.org/asset/48409211 (Cartago,
CR) or up to
14x https://macaulaylibrary.org/asset/48408511 (Cartago, CR)
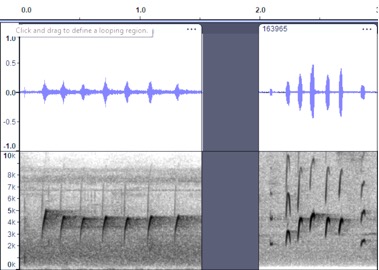
https://macaulaylibrary.org/asset/264163 (Valle de
Cauca, CO) 7x-12; https://macaulaylibrary.org/asset/163965
VOCALIZATION A3 –
Position calls
Taxa: berlepschi
Context: position call,
given commonly, especially right after landing
Description: repeated pip
notes, often trailing off a bit at the end (more spacing between notes,
descending somewhat in pitch). Described by Hilty and Brown (1986) as
“frequently repeats a spirited bubbly ser. of 4-7 pee or pik
notes as flicks tail; call reminiscent of that of Greater Pewee but faster,
weaker, and not as sharp.” [not clear what Greater Pewee vocalization is
referred to here.”] Ridgely and Greenfield (2001) describe as “call an
often-heard fast series of pee or pik notes.”
Discussion:
The
only vocalization from eastern Panama are Cerro Pirre recordings of this call
type (A3) in Macaulay Library with up to 11 notes https://macaulaylibrary.org/asset/25779, up to 22
notes (see below), and on Xeno-canto https://www.xeno-canto.org/360141 with up to 22
and 31 notes in a series. This sounds generally like aurantiiventris,
but the number of notes in each series are markedly longer. Others also sound
similar but also tend to give longer series, such as 12 notes https://www.xeno-canto.org/108571 or up to 24
notes https://www.xeno-canto.org/85120. This
distinction seems to be enough to potentially consider this vocalization to be
different, although the quality of the pip notes is quite similar.
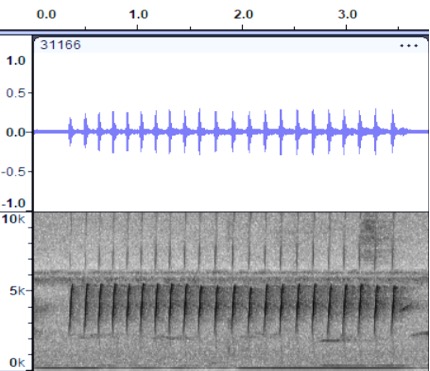
Cerro
Pirre, Panama https://macaulaylibrary.org/asset/31166
VOCALIZATION A4 – Position calls
Taxa: olivaceus
Context: position call,
given commonly, especially right after landing
Description: repeated tup
notes, often trailing off a bit at the end (more spacing between notes,
descending somewhat in pitch)
The
quality of these notes is markedly different from examples in Vocalization A2,
with each note more like a chup or tup than a pip or peep.
These differences seem to equate to species-level differences in vocalizations.

olivaceus, Peru https://macaulaylibrary.org/asset/519282
VOCALIZATION B1 – primary song
Taxa: phaeocercus Group
Context: Primary song
Description: Clear,
high-pitched whistled song, vaguely recalling some Contopus (e.g., Tropical Pewee (Tumbes) Contopus cinereus punensis) (MJI). Howell and Webb (1995) describe
a bright, slightly emphatic seeeu or pseeeu, repeated every few s and mainly
in breeding season.
Discussion: This high,
whistled note appears to be the song, and this is quite similar to the songs of
several pewees. There appear to be subtle but noticeable differences between
all four groups, so these are separated here.
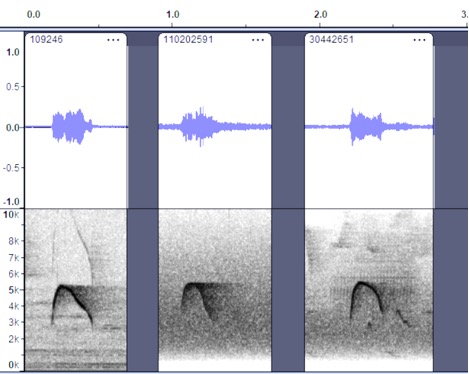
Sinaloa, Mexico https://macaulaylibrary.org/asset/109246; Chiapas,
Mexico https://macaulaylibrary.org/asset/110202591;
Quetzaltenango, Guatemala https://macaulaylibrary.org/asset/30442651
VOCALIZATION B2 – primary song
Taxa: aurantiiventris
Context: Primary song
Description: Clear,
high-pitched whistled song, vaguely recalling some Contopus (e.g., Tropical Pewee (Tumbes) Contopus cinereus punensis) (MJI). Stiles and Skutch (1989)
describe “a thin seer or peew, especially in breeding season”
Discussion: This high,
whistled note appears to be the song, and this is quite similar to the songs of
several pewees. There appear to be subtle but noticeable differences between
all four groups, so these are separated here. In addition to the obvious
differences in the shape of the sonogram, Mexican birds sound more like a peaaar vs. Costa Rican wheeeerr.
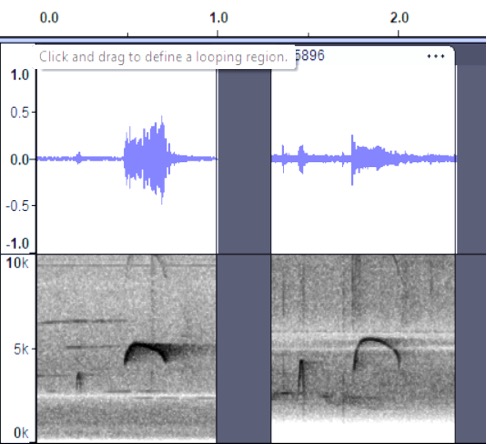
Cartago, Costa Rica, https://macaulaylibrary.org/asset/48408511; Heredia, CR https://macaulaylibrary.org/asset/165896
VOCALIZATION B3 – primary song
Taxa: berlepschi
Context: Primary song
Description: Clear,
high-pitched whistled song, vaguely recalling some Contopus (e.g., Tropical Pewee (Tumbes) Contopus cinereus punensis) (MJI).
Discussion: This high,
whistled note appears to be the song, and this is quite similar to the songs of
several pewees. There appear to be subtle but noticeable differences between
all four groups, so these are separated here. In addition to the obvious
differences in the shape of the sonogram, the song of South American birds
sounds like Sweet! as compared to peaaar
or wheeeerr, for Mexican or Costa Rican birds. It is
too bad that we don’t have a comparable example of this from eastern Panama.
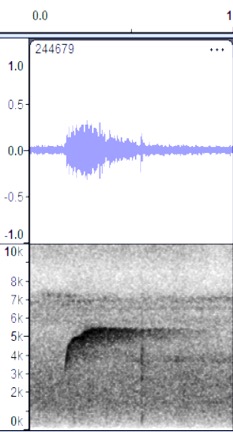
Esmeraldas,
Ecuador https://macaulaylibrary.org/asset/244679
VOCALIZATION B4 – primary song
Taxa: olivaceus
Context: Primary song
Description: Clear,
high-pitched whistled song, descending more than in other taxa and thus quite
different.
Discussion:
Species-specific
and quite distinct high song.
High sieeuu song: Bolivia https://www.xeno-canto.org/44221
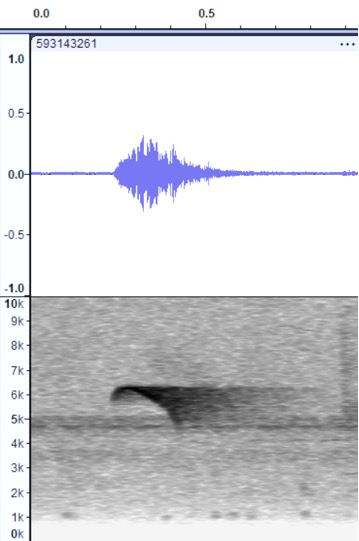
olivaceus, San Martín,
Peru https://macaulaylibrary.org/asset/593143261
VOCALIZATION C1
Taxa: phaeocercus Group
Context: Alternate song; pip-weet song
Description: Similar to the
high, whistled note but with introductory pip notes. This is sort of a
combination of vocalization A (introductory notes) and vocalization B (primary
song). It may serve as a dawn song or a variation on the primary song; note
however that a more complex dawn song is clear in some other examples (see
Vocalization D below). Stiles and Skutch (1989) describe peew-peew or peep-wit
wit, although it is not clear that this is the same vocalization in
examples below.
Discussion: I found no
good examples that equate to berlepschi or olivaceus, but this
taxon does seem to occasionally add a pip note before the song in a
semi-structured way. Here is one example:
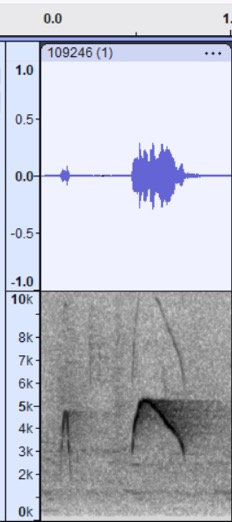
Sinaloa, Mexico https://macaulaylibrary.org/asset/109246
VOCALIZATION C2
Taxa: aurantiiventris
Context: Alternate song; pip-weet song
Description: Similar to the
high, whistled note but with introductory pip notes. This is sort of a
combination of vocalization A (introductory notes) and vocalization B (primary
song). It may serve as a dawn song or a variation on the primary song; note
however that a more complex dawn song is clear in some other examples (see
Vocalization D below). Stiles and Skutch (1989) describe peew-peew or peep-wit
wit, although it is not clear that this is the same vocalization in
examples below.
Discussion: In aurantiiventris,
this may or may not equate to the examples for berlepschi and olivaceus
below, which seem like more structured, more regular, and probably more
commonly given vocalizations. Still, the short pip notes before the song so
match the general structure of the examples C3 and C4.
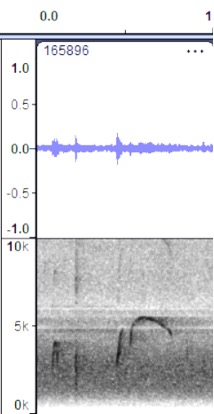
Heredia, Costa Rica https://macaulaylibrary.org/asset/165896
VOCALIZATION C3
Taxa: berlepschi
Context: Alternate song; pip-weet song
Description: Similar to the
high, whistled song but with introductory pip notes. This is sort of a
combination of vocalization A (introductory notes) and vocalization B (primary
song). It may serve as a dawn song or a variation on the primary song; note
however that a more complex dawn song is clear in some other examples (see
Vocalization D below). Ridgely and Greenfield (2001) describe “song a repeated
fast phrase of high, thin notes, e.g., tsu-tsu-tseét, with softer notes
sometimes interspersed between phrases (P. Coopmans).”
Discussion: In berlepschi,
this seems like a well-structured and very intentional song. It is not totally
clear how its significance differs from vocalization B3 and is perhaps just a
variation.
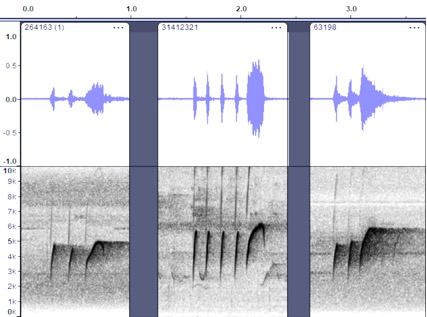
Valle de Cauca,
Colombia https://macaulaylibrary.org/asset/264163; Esmeraldas,
Ecuador https://macaulaylibrary.org/asset/31412321; Esmeraldas,
Ecuador https://macaulaylibrary.org/asset/63198
VOCALIZATION C4
Taxa: olivaceus
Context: Alternate song; pip-weet song
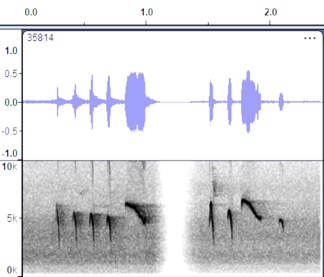
Peru, https://macaulaylibrary.org/asset/35814 (both)
VOCALIZATION D1
Taxa: phaeocercus
Group
Context: Dawn song
Description: pit-whit-ooh-wheer…
pit-whit-ooh-whit-wheer… pit-whit-ooh-wheer
Notes: Quite similar
to Costa Rica recording, with some minor differences in the shape of the notes
and delivery.
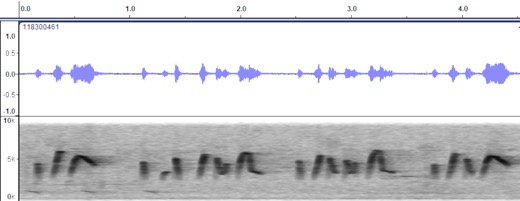
Sololá,
Guatemala https://macaulaylibrary.org/asset/118300461
VOCALIZATION D2
Taxa: aurantiiventris
Context: Dawn song
Description: pip-pip-pip-pip-pip-pip-WHEEooo
pip-pip-pip-pip-wheEEE
Notes: Markedly
slower delivery in Costa Rican birds compared to Colombia recording. Stiles and
Skutch (1989) describe quaint, prolonged dawn song a very rapid series of high,
thin notes: bip-bip-bip-dididi-up-bip-bip-bibibiseer
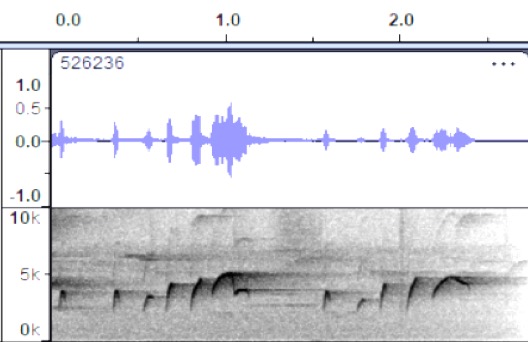
https://macaulaylibrary.org/asset/526236
VOCALIZATION D3
Taxa: berlepschi
Context: Dawn song
Description: pi-pit
pi-pi-pit pi-wheoo….pi-pit wheer…
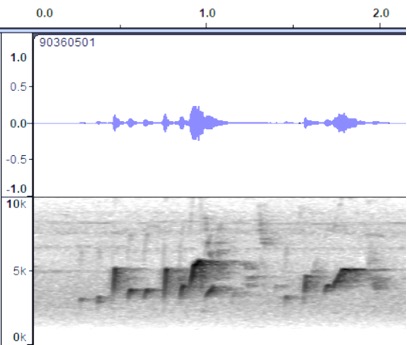
Valle de Cauca, Colombia https://macaulaylibrary.org/asset/90360501
VOCALIZATION E
Taxa: all subspecies
groups (except no examples found for berlepschi)
Context: short contact
call
Description: Short, abrupt
"pip", recalling Alder Flycatcher Empidonax alnorum (MJI). Howell
and Webb (1995): a sharp, usually
quiet pic or beek, suggesting Hammond's Flycatcher
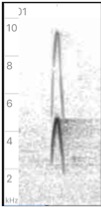
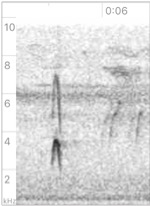

Left:
phaeocercus Group, https://macaulaylibrary.org/asset/524464 (Sinaloa, MX).
Multiple examples on this cut vary in strength and are sometimes given in
advance of Vocalization D (see below). See also https://macaulaylibrary.org/asset/216981; center: aurantiiventris,
https://macaulaylibrary.org/asset/72888 (Costa Rica);
right: olivaceus, https://macaulaylibrary.org/asset/35814 (Peru).
Probably
sounds fuller and lower in olivaceus and maybe diagnosably different
from the other taxa. I did not find examples for berlepschi, so it is
unclear if they have a similar note, but it seems likely. A single note given
in https://xeno-canto.org/85120
at 0:11 is likely analogous to this note. Given that the strength
and stridency of this note varies according to how excited the individual is, I
did not detect obvious differences between the taxa in this note. They may
exist, but it would require a larger sample to confirm this. To my ear, in the
examples I heard, this note seems similar across the genus, even between the
two taxa currently treated as separate species (phaeocercus sensu
lato and olivaceus).
VOCALIZATION F
Taxa: Costa Rican aurantiiventris only [likely all?]
Context: aggression
calls
Description: kew…kew

https://macaulaylibrary.org/asset/48408511
VOCALIZATION
G
Taxa: Costa Rican aurantiiventris only [likely all?]
Context: aggression
calls
Description: kew…kew
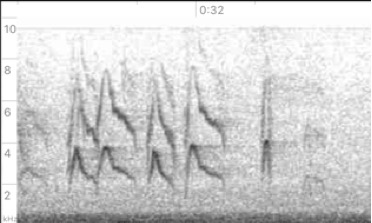
note "pip"
at end
https://macaulaylibrary.org/asset/48409211
Marshall
Iliff and Pamela C. Rasmussen, May 2026
Vote
tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Stiles: “A very
interesting exposition of the variation within the M. phaeocercus group,
which clearly merits splitting into 3-4 species. Most of this variation in
centered in NACC territory, and the options for recognition by SACC boil down to
two, labeled options A and B, which differ in treating the South American form berlepschi
as a species, or as a subspecies of the taxon aurantiiventris (which
occurs from Costa Rica southward). The two options given for SACC are to
consider M. phaeocephalus as consisting of three species (A:
nominate phaeocephalus, aurantiiventris (Costa Rica and western
Panama) and berlepschi (eastern Panama to northwestern Ecuador), or B:
two species, phaeocephalus and aurantiiventris with berlepschi
as a subspecies, thus including Costa Rica through NW Ecuador).
“My vote
is YES for option A (three species). I consider that berlepschi differs from aurantiiventris
in most of its vocal repertoire, plumage color and pattern, biogeography
(habitat, elevation) to an extent comparable to the species-level differences
in several related genera of tyrannids (genetic data are incomplete but do not
conflict with this interpretation). Regarding E-names, I agree that all Mitrephanes
taxa are distinctive enough within the family to justify the group-name of
Tufted-Flycatcher, and that a suitable name for berlepschi would be
Chocó Tufted-Flycatcher.”
Comments from Lane: “1. YES, tentatively, to option A. With only a
cursory listen, I hear only weak differentiation between voices of aurantiiventris
and berlepschi, but I suspect if I really studied the two, I would be
more impressed by their differences. Plumage-wise, they seem pretty
distinctive, and it is clear to me that M. phaeocercus is truly the
vocal outlier of the complex, with M. olivaceus being an outlier in
having no obvious dawn song (I’ve certainly never detected one, and it’s a bird
I have encountered a fair amount… though rarely at dawn, admittedly).”
Comments
from Robbins:
“1. Option
B. Vocalizations,
plumage, and genetic data clearly support recognizing two species, phaeocercus
and aurantiiventris.
“With
regard to whether berlepschi should be treated as separate from aurantiiventris,
I feel the data at hand are inconclusive. As pointed out, the Harvey (2020)
study lacked genetic sampling for aurantiiventris. With regard to what has been presented for
vocal differences between these two taxa, it is stated that the primary song
between these two is different. However, based on the single examples, like
Dan, I find that they are similar. With regard to dawn song, again based on
single examples, when one clicks on the example for aurantiiventris the
message comes up ‘restricted access’. I
did a filtered search (dawn song) for Tufted Flycatcher and that example
(#526236) doesn’t come up. Clearly, the
plumage is distinct between these two. Thus, what I would like to see before
supporting the split of aurantiiventris and berlepschi is at
least a more robust presentation of vocal differences, i.e., multiple examples
that are clearly identified and made accessible.”
Additional
comments from Robbins:
“Following
my comments on this proposal I looked at quite a few photos, here are two that
left me puzzled concerning berlepschi.
“https://ebird.org/checklist/S87205736
“https://ebird.org/checklist/S150437416”
Comments
from Areta: “The most
important line of evidence in this case are the vocalizations, and the limited
genetic sampling. These argue for a clear split separating M. phaeocercus including tenuirostris
from M. aurantiiventris
including berlepschi
and a more muddied situation to further split berlepschi from aurantiiventris.
“Position
calls:
There is considerable variation in the "position calls" within the phaeocercus group, including
short and rapid calls with little burry quality and long, slow calls which
sound quite burry. Given that there are few samples, it is difficult to assess
whether variation is geographic or purely individual or both. But the bottomline is that there is a lot of variation. Despite
variation, the phaeocercus
group differs markedly from the aurantiiventris
group. Once variation has been acknowledged within the phaeocercus group, focusing
on the aurantiiventris
group becomes interesting: while it certainly seems that aurantiiventris has a more
musical quality, and perhaps more clearly "bent" and inflected notes,
there is quite a lot of variation in both aspects in berlepschi calls, both
regarding rate of note delivery and note shape. Looking and hearing at the
spectrograms, I cannot discard clinality across the geography. Of course,
clinal variation need not be the same across space, and having a small leap in
variation does not immediately equates to having two different species.
“Please
note that in Figure 5 the names berlepschi
and olivaceus have
been swapped (the one on the right is olivaceus,
the one on the left is berlepschi):
the calls of olivaceus are radically different, while the calls of berlepschi and aurantiiventris bear a
strong resemblance.
“Dawn
song:
again, the break between the phaeocercus
and aurantiiventris
groups seem outstanding (despite reduced sample sizes: how many recordings were
there available?), while the differences within taxa in the latter are less
compelling or difficult to interpret without adequate sample sizes to test for
variation between individuals vs. geographic variation: although it seems that berlepschi is a more rapid
singer with shorter notes than aurantiiventris,
this requires more evidence. Until then, the evidence seems limited. If someone
finds a decent series of dawn songs that can be compared, I´d like to hear
about that.
“Pip
calls:
these are tough to distinguish in general, also in Contopus. So, I wouldn´t place too much hope on
these. But if the differences are obvious, then they do support species-level
divergence.
“Plumage
seems not to perfectly distinguish aurantiiventris
and berlepschi,
with people in NACC and SACC commenting on this.
“With
all this in mind and given the lack of formal acoustic analyses and no
molecular phylogenetic information for aurantiiventris,
I am comfortable with only
splitting M.
phaeocercus
including tenuirostris from M. aurantiiventris including berlepschi at this point
[i.e. NO to Option A, YES to Option B].”
“B. I am fine with using "Tufted Flycatcher" or
"Tufted-Flycatcher" for Mitrephanes] (I would also be fine
with just using Mitrephanes as a common name, a line of thought that I
like more each day). Before discussing the E names, let´s see what proposal
passes. But preliminarily, I endorse Cinnamon or Pine-forest “Tufted-Flycatcher"
for M. phaeocercus and Yellow-bellied or something?-for’est
"Tufted-Flycatcher" for M. aurantiirostris (including berlepschi).
If the three-species treatment gains traction, I may lean towards the
habitat-based names, including Pine-forest, Cloud-forest and Choco.
Comments
from Glenn Seeholzer (guest voter):
“1. YES,
to Option B. phaeocercus/tenuirostris should clearly be split from
aurantiiventris + berlepschi based on dramatically different vocalizations,
the position notes in particular, and plumage differences. Following Nacho's
logic, I shifted my focus to aurantiiventris and berlepschi and
found insufficient qualitative evidence to support splitting these two taxa
(Option A). Option B is a clear improvement over the current arrangement and
the more prudent choice based on the qualitative evidence available. Rationale
below.
I was not
particularly impressed by the vocal differences between aurantiiventris
and berlepschi but note that they are at least as different as either
are from olivaceus. I expect that a quantitative analysis of at least
the 'position' notes among the four taxa would result in aurantiiventris
+ berlepschi + olivaceus being highly clustered and probably
overlapping, with phaeocercus/tenuirostris separated off to the
side. Nonetheless, because genetic and plumage data clearly support the species
status of olivaceus, one could argue that aurantiiventris and berlepschi
should be considered species as well in spite of their subtle vocal
differences.
That said,
before considering aurantiiventris and berlepschi a distinct
species, I think a quantitative analysis of the entire group is needed. First,
we have a potentially new taxon from Cerro Tacarcuna. Second, I quickly
examined the geographic variation in vocalizations within phaeocercus/tenuirostris
(see galleries below) and feel they do not align with the described ranges of
these two taxa. The vocal break in their position notes and dawn songs seem to occur
across the Isthmus of Tehuantepec, rather than across the transition from the
described ranges of tenuirostris to phaeocercus (somewhere around
Michoacán). I know Central American biogeography well enough to say that a
break across the Isthmus of Tehuantepec is= not surprising, but is there
precedent for a biogeographic break around Michoacán? Regardless, the vocal
differences across the Isthmus of Tehuantepec are at least as distinctive as
the differences in the position notes between aurantiiventris, berlepschi,
and olivaceus. Could there actually be six species involved? Seems like
a quantitative treatment is the only way to sort these remaining questions out.
#####
Below are
audio galleries of phaeocercus/tenuirostris. Group A and Group B
have largely the same position notes and dawn songs with a sharp break in these
same vocs across the Isthmus of Tehuantepec to Group
C.
Group A. 'tenuirostris'
(western Mexico, Jalisco to Arizona):
Group B. 'phaeocercus'
(east from Michoacán to west of Isthmus of Tehuantepec):
“Group C. 'phaeocercus' east of Isthmus of Tehuantepec (Chiapas, Guatemala,
Honduras):
“YES to use the group name ‘Tufted-Flycatcher’
for all members of Mitrephanes
Comments
from Remsen
“YES for
Option B – two species. The case for treating the phaeocercus group from the aurantiiventris is solid based on vocalizations,
although I would prefer that all these valuable details had been published in a
separate paper (and would still encourage Marshall and Pam to publish all this
and add Glenn as a co-author to include all his analyses, so that all their
hard work reaches its full potential; they’ve already done most of the hard
work).
“The
case for three species is weaker, as articulated by Dan, Nacho, Mark, and
Glenn, and requires additional data and analyses. If this proposal were
published as a paper, then those needs could be specified and attention drawn
to them.
“YES
for using Tufted-Flycatcher as a group name. I agree with all the points about
the need to replace as many “Something Flycatcher” names with group names when
possible.”