Proposal (1077) to South
American Classification Committee
Treat Lampropsar tanagrinus as consisting
of three species
Background: SACC currently treats Lampropsar tanagrinus (Icteridae) as a
single species with the following footnote:
24b.
Schultz et al. (2026) used genetic and morphological evidence to advocate
treating the subspecies guianensis and violaceus as separate
species. SACC
proposal badly needed.
Dickinson and Christidis (2014) treated the species as consisting
of five subspecies:
L. t. guianensis of
the Guianan Shield
L. t. tanagrinus of
the central and western Amazon
L. t. macropterus from
upper Juruá
L. t. boliviensis of
northeastern Bolivia
L. t. violaceus of
Mato Grosso
They placed the first three in a single group, and the last two
each in their own group, citing Sibley & Monroe (1990). They also cited Jaramillo-Burke (Icteridae
book) and Fraga (HBW chapter) for opinions that more than one species may be
involved.
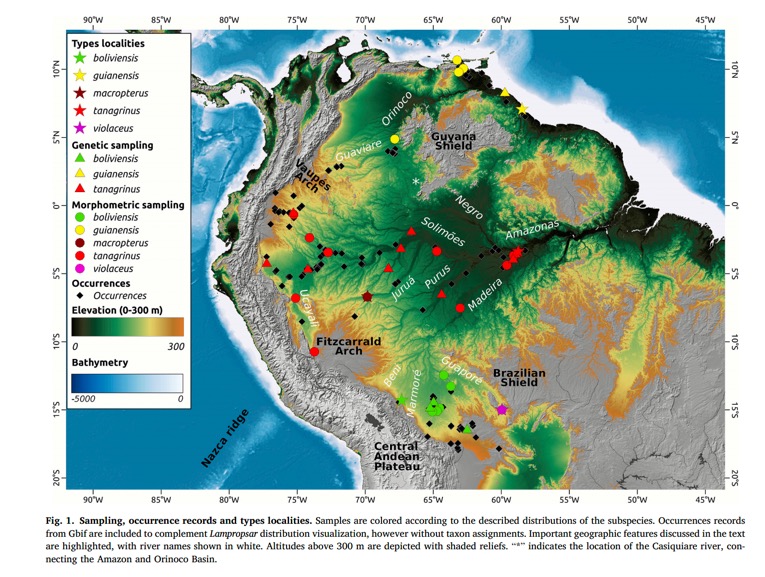
New information: Schultz et al. (2026) analyzed
genetic (mtDNA and UCEs) and morphometric variation (bill length, bill width,
wing length, tail length, tarsus length) in Lampropsar tanagrinus. See map below
for sampling distribution (which is, incidentally, a great one in my opinion
for understanding Amazonian biogeography, especially with respect to
floodplains).
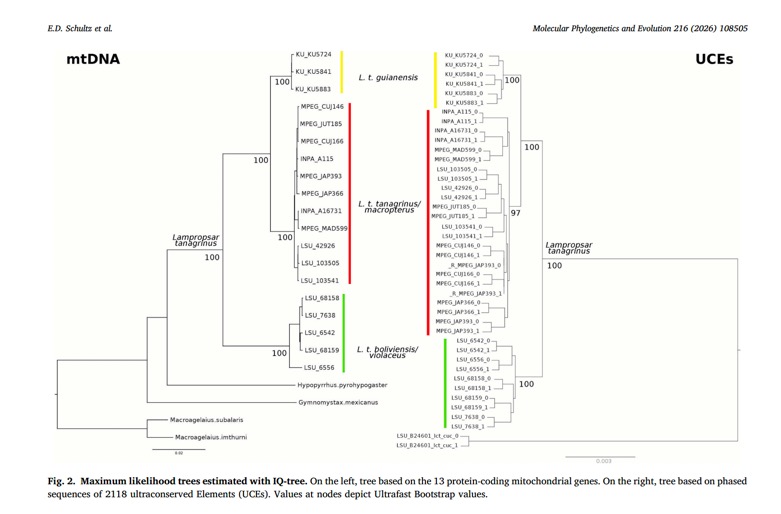
The findings with respect to species limits is that the mtDNA and
UCE results both showed three strongly supported, separate groups:
The morphometrics were presented as boxplots:
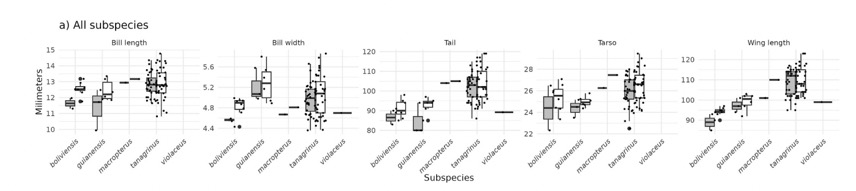
My takeaway from this is that nominate tanagrinus is
notably larger in wing length and tail length than either guianensis or boliviensis,
with little or no overlap, and that guianensis is also larger than boliviensis
in wing length, with minimal overlap. I
predict that if all 5 measurements were plotted in multidimensional
morphospace, sex by sex, that the clouds might not overlap.
Based on the genetic data showing three distinct lineages, and to
some extent the mensural data, the authors argued to species rank in an
implicit evolutionary species concept framework.
Discussion: I can’t be entirely objective on
this one because a specimen of boliviensis that I collected is what
first brought me to LSU in 1977 because I was certain from previous in my
experience with Lampropsar tanagrinus in the Leticia region that
whatever I had collected in eastern Bolivia in 1977 was not the same species as
L. tanagrinus; in fact, it even didn’t occur to me at the time that the
bird I collected, which turned out to be boliviensis [=violaceus]
was a Lampropsar Long story,
worth hearing in full in another venue, but meanwhile see my Ebird list (https://ebird.org/checklist/S14691582)
for part of the longer story. So, for 50
years, I’ve been convinced that boliviensis deserved species rank and
eagerly read the paper hoping for slam dunk evidence.
Although all the data in Schultz et al. are consistent with
species rank for all three groups, unfortunately, I do not think they are
sufficient for a change, and here is why:
1. No vocal data.
Differences in voice were how I “knew” boliviensis was different, but
that was based on my superficial knowledge of, and ex-post-facto comparisons of,
vocalizations that I could not be sure were homologous, influenced also by an
almost certain eagerness to have found evidence for species rank. Differences
in voice are associated with breaks in gene flow in closely related icterids,
e.g., Quiscalus major vs. Q. mexicanus here in the USA, Agelaius
phoeniceus-A. assimilis-A. tricolor but not A. p. bicolor,
and lack of documented differences in voice is a primary reason we voted to
keep armenti as a subspecies of Molothrus aeneus. Etc. So, to be on solid footing for a
species-level split, I think vocal analyses are necessary. In fact, I think their absence is sufficient
reason to reject the proposal, regardless of the other points below. I suspect that one could do a “dry lab”
analysis of online recordings to address this, with a short paper with the
sonograms. So, I don’t think it’s asking
for much to analyze vocalizations, which have been a mainstay of species
delimitation in the Icteridae starting with Bud Lanyon’s seminal work on Sturnella
6-7 decades ago. Also, Jaramillo and Burke (Icteridae book, 1999) mentioned
possible vocal differences in violaceus, so it would be nice to clear
that up.
2. Sampling gap. Notice the
substantial gap in sampling in the area in Fig. 1 above in eastern
Colombia-Venezuela between the tanagrinus and guianensis genetic
samples. In terms of known occurrence,
the two taxa actually approach each other there – see the black triangles in
“Vaupes Arch” and “Guaviare” area of the map.
From topography, I suspect that area is occupied by guianensis,
which “ought to be” separated from Amazonian tanagrinus by the Vaupes
Arch. Also note there is but a single sample (in the middle Orinoco) in their
analysis away from the coastal populations of guianensis. I’m likely being overly picky on this point,
but technically without sampling in that gap, which looks to be about 800 km
long, one can’t officially rule out the existence of a cline. I doubt that’s the case, but I’m queasy about
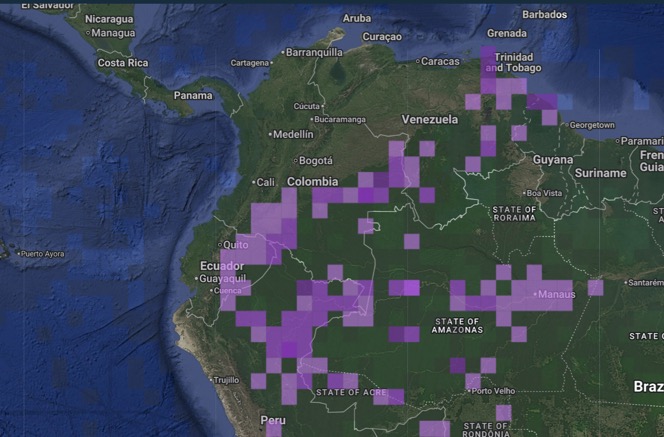
the absence of information from anywhere near the contact zone. Here’s the eBird map, which makes the
distribution of tanagrinus and guianensis appear continuous in
eastern Colombia:
Also, the degree of genetic divergence is not impressive. This doesn’t make any difference to me personally
in species delimitation, but it does to some of you. Guianensis was estimated to have
diverged from nominate tanagrinus only ca. 800K years ago, and they from
boliviensis only ca. 2.75 MYA.
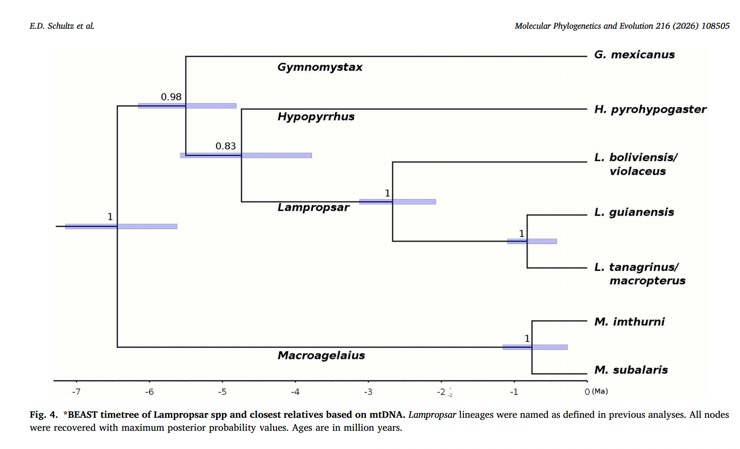
Not enough taxa are included to place these relative distances in
a valid comparative framework.
An analysis of plumage would also contribute to the analysis of
species limit in Lampropsar because three of the named species used
plumage differences in their descriptions. Also, Blake in Peters 1968 stated
that boliviensis intergrades with violaceus in Santa Cruz,
Bolivia, presumably based on plumage.
However, Schultz et al. examined the type specimen of violaceus
and did not detect any differences in its plumage, which was the basis for
Hellmayr’s description of the taxon in comparison to nominate tanagrinus
(“easily distinguished by the violet or purplish luster of the whole body [of violaceus]”)
but not boliviensis, which was not described at the time. It would be nice to clear all this up.
Tidbits:
Schultz et al. found that L. t. macropterus is just a point
on an east-west cline of wing length and is embedded in the tanagrinus
genetic cluster; hence, this taxon should be treated as a synonym of nominate tanagrinus.
They also examined the holotype (and evidently only) specimen of violaceus
and found no differences between it and boliviensis specimens; violaceus
Hellmayr 1906 has priority over boliviensis Gyldenstolpe 1941. The
type locality of violaceus is at the edge of the Pantanal and is
therefore isolated from the Beni-Mamoré (boliviensis) by the Chiquitano
dry forest uplands and is in a different drainage system, so I think it might
be worth taking a second look at that specimen with colorimetry.
Schultz et al. make the standard boiler plate plea for ranking
separate evolutionary lineages as species and preach that the BSC masks
diversity, but they do not point out that the subspecies unit, which they used
to frame their analyses, can also be used in any analyses of diversity, of
evolutionary patterns, and of conservation priorities. Subspecies taxa,
properly delineated, are a level of biodiversity
Recommendation:
I recommend a NO on this species-level split for the reasons
articulated above. I do not think that the evidence in Schultz et al. has quite
reached the point of forcing burden-of-proof on maintaining the single species
classification. I hope that someone undertakes the necessary analysis of
vocalizations, because I suspect that more than one species is involved. Schultz et al. have established a firm
foundation for further research.
References:
SCHULTZ, E. D., J. CRACRAFT, M. FERREIRA, AND C. C. RIBAS. 2026.
Deep diversification in Lampropsar tanagrinus (Aves: Icteridae)
illustrates hidden diversity and its implication to biogeographic inferences in
the Neotropics. Molecular Phylogenetics
and Evolution 216: 108505.
Van Remsen,
June 2026
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments from
Robbins: “NO. Although the UCE data
clearly indicate three groups, I agree with Van's comments about the lack of a
vocal analysis. I would like to see that before splitting Lampropsar
tanagrinus into multiple species.”
Comments from
Lane: “Perhaps this proposal should be
set up as “A) split violaceus from the remainder of the complex
(Yes/No), B) split guianensis from tanagrinus, regardless of vote
in A (Yes/No).” If constructed this way, I vote: A) Yes, B) No. I think the
evidence provided, and from a quick perusal of sound recordings, that we can at
least split L. violaceus/bolivianus from the remainder of L.
tanagrinus (incl guianensis) and then can deal with the latter
complex in the future.”
Response from
Remsen: “That might have been a better
way to set it up, but there were no vocal data in the original paper, and it is
not SACC’s responsibility to do research, only to vote on what has been
published. That doesn’t mean that we
can’t supplement points with our own research, e.g., adding recordings, but I
don’t think we have enough resources to introduce entire new data sets
including proper analyses of them, and a few samples would be insufficient in
my opinion. So, if the proposal fails,
then I hope someone will take on the task of a fully developed analysis of
voices. In fact, I really hope that
happens because I would like to have my assessment from 50 years ago affirmed
by actual data.”
Comments from Areta: “I am torn by this case.
It is clear that there are three clades, but I don´t see any other reason
(morphological, vocal, ecological) to afford these clades species status under
the BSC and allied concepts such as the RSC. The failure of the work to provide
“diagnostic diagnoses” only adds to my worries.
“The yardstick approach would seem to indicate that
the level of genetic differentiation between guianensis and tanagrinus
(including macropterus as part of a cline) are comparable to those
between Macroagelaius imthurni and M. subalaris, but this seems
to be all we have to decide on their taxonomy. The greater degree of
differentiation of violaceus/boliviensis adds slightly more support for
this split.
“I vote NO to any split for the time being, although
with much regret over the lack of further biological evidence to interpret the
genetic differences.”