Proposal (1079) to South
American Classification Committee
Species limits in Myrmotherula
longipennis: A. Treat three subspecies groups as separate species;
B. Establish English names for the three species.
Background: The current
SACC note is as follows:
24. Ridgely & Tudor (1994)
and Zimmer & Isler (2003) suggested that Myrmotherula longipennis
may consist of more than one species; the subspecies garbei was treated
as a separate species from Myrmotherula longipennis by Cory &
Hellmayr (1924), but they were treated as conspecific
by Peters (1951). Isler et al.
(2025) provided rationale for treatment of Myrmotherula longipennis as
three separate species: (a) M. longipennis of northern Amazonia, (b) M.
garbei, including subspecies zimmeri, of southwestern Amazonia, and
(c) M. paraensis, including subspecies ochrogyna and transitiva, of southeastern Amazonia. SACC proposal badly
needed.
The
current classification is as follows (Zimmer and Isler 2003 HBW, Dickinson and
Christidis 2014 H&M): The subspecies are described mostly on the basis of
plumage color.
M. l. longipennis: ne. Ecuador N
of the Napo east to Guianans and ne. Brazil
M. l. zimmeri: e. Ecuador
and ne, Peru S of the Napo and N of the Marañón
M. l. garbei: e. Peru and
n. Bolivia S of Marañón-Amazon and east to w. Brazil to Rio Madeira
M. l. ochrogyna: S of the
Amazon in Brazil from R. Madeira east to R. Tapajós
M. l. paraensis: S of the
Amazon in Brazil from R. Tapajós to Maranhão
M. l. transitiva: s. Amazonian Brazil in upper Madeira and
Tapajós
Here
are some specimens.

New
information:
Isler et al. (2025) analyzed vocal differences with respect to five types of
vocalizations as well as plumage color differences from throughout the
distribution of M. longipennis, with samples from all named
populations. In a companion paper,
Chesser et al. (2025) analyzed genetic differences from throughout the range.
Sampling
units
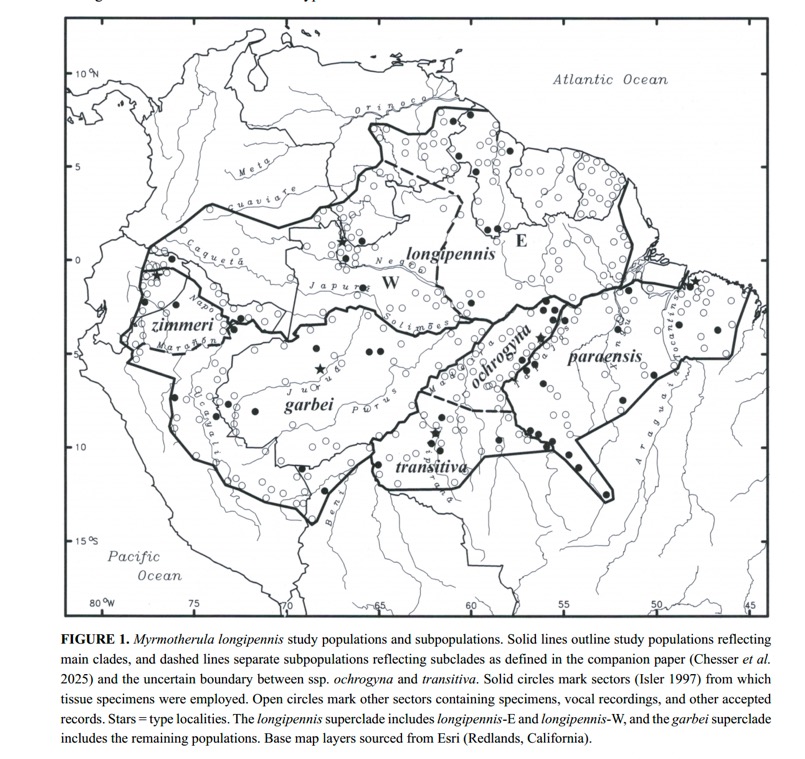
They
grouped their samples according to the boundaries of the described subspecies,
as in the map below. An additional partition was used in the analyses: western
and eastern populations of nominate longipennis because these formed
separate clades in Chesser et al. (2025). Note that there do not appear to be
any useful contact zones between the three proposed species, and the rivers
delimit all taxon boundaries except transitiva
and ochrogyna.
Vocalizations
Isler
et al. (2025) assembled a library of 501 recordings of Myrmotherula
longipennis to analyze geographic variation in of five separate
vocalizations the species geographic range. Here is a snip from their Methods:
“Criteria for diagnosability of vocal characters followed methods
developed in an earlier study (Isler et al. 1998). For quantitative characters,
we required the absence of range overlap and the likelihood of non-overlap with
larger sample sizes. For qualitative characters, such as note shape, we
required consistent visual diagnosability as determined in blind tests. We
employed quantitative and qualitative characters for songs, but only
qualitative characters for calls. We identified quantitative song characters in
stages. In the initial stage, we plotted the locations of recordings on digital
maps, and we selected three recordings of songs from different parts of each
study population’s geographic range on the basis of recording quality. We
obtained twenty-six measurements from spectrograms of songs and computed 19
ratios (e.g., first note length/terminal note length) from these measurements.
We obtained measurements from spectrograms projected on a 43-cm screen with the
default settings of RAVEN Pro 1.4 (Charif et al.
2010), except that the display was set to smooth, overlap was adjusted from 50
to 93.7% depending on the recording’s quality, and contrast was adjusted
according to the recording’s intensity, with care taken to retain all elements
of the vocalization.
And:
“We then expanded the analysis to include all available and usable
recordings (except that we sampled randomly from locations for which there were
20 or more) and again eliminated characters whose ranges of measurements overlapped
among all pairs of study populations (except in rare instances in large samples
where extraordinary values were considered anomalies).”
Then,
they analyzed the recordings to search for and test for diagnosable characters
– see their paper for details.
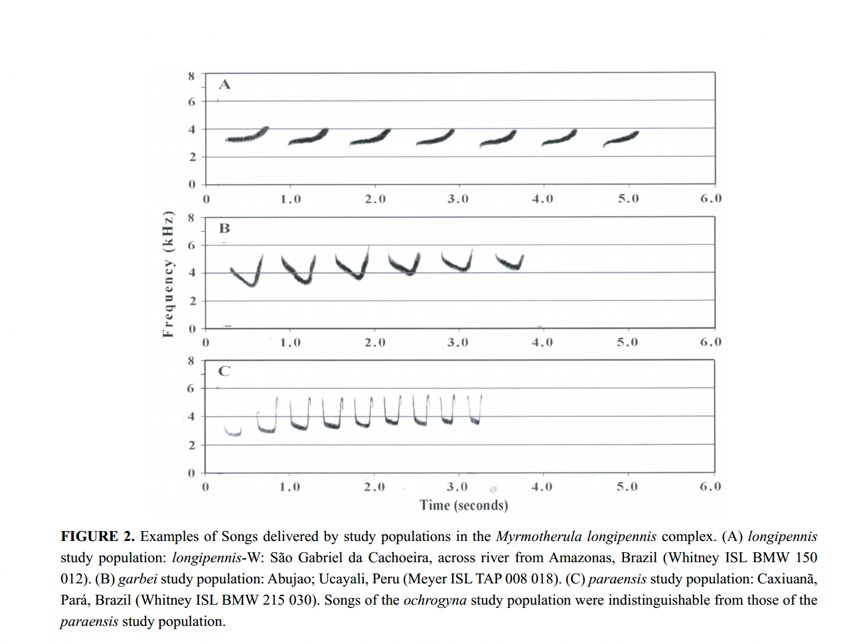
For
loudsongs, they found three song types that corresponded to (1) nominate longipennis,
(2) garbei, and (3) paraensis + ochrogyna. In addition to note
shape, see the difference in pacing between paraensis + ochrogyna vs.
the other two:
Examples:
•
longipennis:
https://xeno-canto.org/1056488 (Dante
Buzzetti)
•
garbei (zimmeri):
https://xeno-canto.org/61377 (by Andrew
Spencer)
•
garbei:
https://xeno-canto.org/688235 (by Jacob
Wijpkema)
•
paraensis:
https://xeno-canto.org/753417 (by Mateus
Gonçalves Santos)
They
also found that the “Series Call” of nominate longipennis differed
diagnostically in note shape from any of the other populations. Also, the “Multi-note
Calls” of the longipennis and garbei study populations differed
diagnostically in note shape from those of the ochrogyna and paraensis
study populations.
Plumage
color
Isler
et al. (2025) also obtained reflectance values (using spectrophotometer) for
four body regions for 76 females; these were the regions used in the
descriptions of the subspecies. They also examined plumage variation in males
and found that all differences were clinal and thus were not used in taxonomic
conclusions. Reflectance values in
female plumage proved to diagnose all six subspecies with 100% accuracy in
discriminant analysis except misclassifications between paraensis and garbei
of 5 specimens.
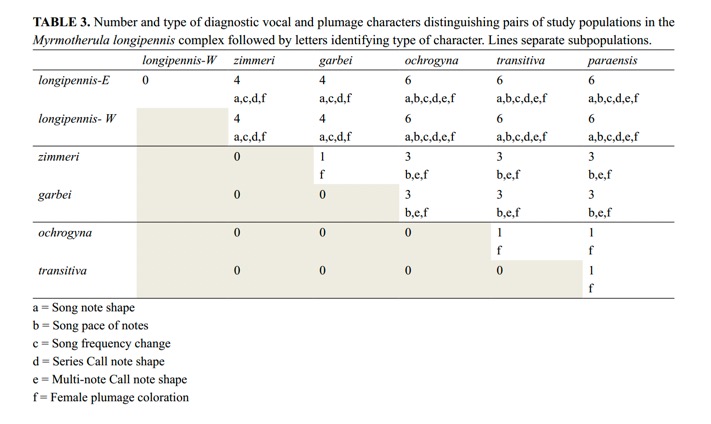
Isler
et al, provided a summary table of the differences between pairs of taxa:
Genetic
data
Chesser
et al. (2025) used DNA sequence data (ND2 and ND3 mitochondrial genes) to
construct the following phylogenetic hypothesis:
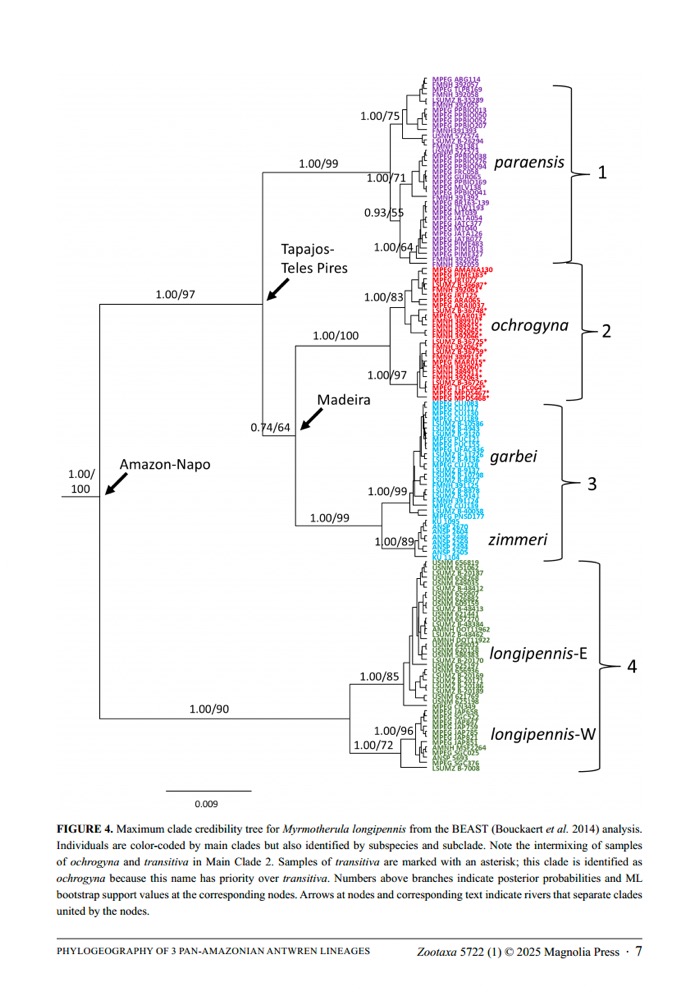
Of
interest is that mtDNA identified two lineages within nominate longipennis
that are fairly differentiated from each other, but that the two lineages have
no known phenotypic differences.
PART
A
Taxonomic
Recommendations
Isler
et al. (2025) used the following rational for ranking taxa as species or
subspecies:
“We recommended species status under the Biological Species Concept for
populations that differed diagnostically in vocalizations (Johnson et al.
1999). Based on a study of vocalizations of syntopic, congeneric species-pairs
of antbirds (Isler et al. 1998) and subsequent applications, vocal differences
were considered diagnostic at the species level if three or more independent
vocal characters were identified. A finding of three distinct characters was
not viewed as a requirement but a guideline, however, and two vocal characters
were considered acceptable for populations that were clearly differentiated by
plumage based on spectrophotometric measurements or on distinct plumage
features. Plumage descriptions in taxonomic recommendations include currently
recognized characters. Study populations that differed in one or two characters
and did not meet requirements for recommendation as species were classified as
subspecies.”
Implementing
these criteria, they recommend the following three-species revision:
Myrmotherula
longipennis
for nominate longipennis
Myrmotherula
garbei,
including also zimmeri as a subspecies.
Myrmotherula
paraensis,
including also transitiva and ochrogyna
as subspecies
Discussion
and recommendation: Iser et al. integrated data from vocalizations, female
plumage, and genetic lineages to produce their recommended classification (and
provided useful diagnoses for all taxa). But for me, the song differences alone
with sufficient and necessary conditions for placing burden of proof on
anything but a three species classification. The additional data are
biologically important and interesting but none on their own would be sufficient
for species recognition. I recommend a YES for following their taxonomic
recommendations.
A
potential reason to vote NO would be that the evidence for treating nominate longipennis
from all the rest is stronger quantitatively and qualitatively than the
evidence for garbei and paraensis being treated as separate from
each other; however, the songs of the latter two differ strongly in pace, which
is a good indicator of separate species rank in avian vocalizations.
PART
B
English
names:
Isler et al. recommended adding directional modifiers to the three species
names as follows: Northern Long-winged Antwren, Western Long-winged Antwren,
and Eastern Long-winged Antwren.
Although I would normally ask for a separate proposal on English names,
this is one of those cases that I see no hope of improving on what Isler et al.
recommend. The plumage differences among
the females are slight, and the vocal differences don’t seem (to me) amenable
to descriptive names. Retaining “Long-winged”
(itself a pretty useless name) as a group name makes these allospecies stand
out as a unit amid the lengthy list of species called Antwren and retains the
link to the parental specoes, and the directional
names, albeit duller than dirt, function to place them in a geographic context.
So, also indicate in your comments whether we should just go ahead and adopt
these names or wait for a separate proposal (that you are willing to write)
with inspired names. I also suspect that if there were more appealing names out
there, then that author team would have come up with them. (Cory & Hellmayr
1924 is of no help because only 2 of 6 taxa were described by then.
For
voting purposes, a YES means you favor adopting the names proposed by Isler et
al., and a NO means that you favor some other option for which you are willing
to write a quick proposal if Part A (species limits) passes. If you vote NO on
Part A, you can either abstain or still vote either way on Part B.
References (if anyone
needs pdfs, just let me know):
CHESSER, R. T., M. L. ISLER, A.
SANTANA, E. K. LATCH, K. F. STRYJEWSKI, J. REED, L. NAKA, R. C. FLEISCHER, AND
A. ALEIXO. 2025. Comparative phylogeographic patterns in three
pan-Amazonian antwren lineages (Aves: Passeriformes: Thamnophilidae: Myrmotherula and Isleria). Zootaxa
5722: 1–44.
ISLER, M. L., R. T. CHESSER, K. F. STRYJEWSKI, AND B. M. WHITNEY. 2025.
Systematics of three pan-Amazonian antwren lineages (Aves:
Passeriformes: Thamnophilidae: Myrmotherula and Isleria). Zootaxa 5722: 45–78.
Van Remsen,
July 2026
_________________________________________________________________________________________________
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments
from Robbins: “YES. First, as has come to be expected, Isler et al. have done
a herculean effort in their analysis of the longipennis group.
Extraordinary that they assessed over 500
vocalizations! Thus, I have no doubt that their recommendations are spot on.
For my own edification (I was unfamiliar with paraensis), I listened to
quite a few vocalizations on Macaulay and the examples provided in the proposal
are indicative of the loud song of the proposed three species split. The
mitochondrial data may be misleading (note poor node support) with regard to
relationships outside longipennis (sensu stricto).
“So, I vote for recognizing three
species as proposed by Isler et al.”
Comments
from Lane: I think
the excellent Isler study can be accepted at face value, and their
recommendations in a 3-way split seem appropriate. Vocally and in (female)
plumage, these are distinctive birds that have been patiently waiting for this
split to happen, so I am happy to oblige. A) YES to the 3-way split. B) not
strongly compelling names, but I think this genus will be another that will
have more splits (M. menetriesii, M. axillaris, etc.) that will
make the creation of more memorable English names very difficult, so this may
be our best hope, so YES.”
Comments
from Andrew Spencer (voting for Claramunt):
“Part B -
YES to the proposed compound names. I tried without avail to come up with
anything better, but as Van says, if there were better names to be had the
authors would have mentioned them. There are many names that one could coin
that would be accurate, but all of those names would be equally accurate to
others in the genus. So, I don't think it's a good idea to try to force a
"more compelling" name that would also probably cause more
confusion.
“I would
also add that in general I'm not opposed to long compound names in these types
of splits. In the field many birders will probably just use the shorthand
"Long-winged Antwren" to refer to them, and then hopefully know which
exact species is the one in question by their location. These names also have
an advantage of keeping existing field guides a bit more relevant, with the
identification features between the various split species and other Myrmotherula
still relevant and easier to find with the link to the pre-split name.”