Proposal (461a, b) to South American Classification Committee
Remove Busarellus
from buteonine genera in linear sequence of Accipitridae and rearrange linear
sequence on non-buteonine genera
Part a: Move Busarellus to follow Ictinia.
Effect on South American CL:
This proposal would overhaul our linear sequence of accipitrid genera.
Background
& New Information & Analysis:
The genus Busarellus has
traditionally placed within the buteonine genera of the Accipitridae, but this
is clearly not correct. The relevant Note from our SACC classification is:
17a.
Olson (1982) found morphological evidence that Busarellus may be more
closely related to a group of largely Old World genera (Milvus, Haliastur,
Haliaetus, Ichthyophaga) than to the New World genera with which
is traditionally associated in linear sequences (e.g., Friedmann 1950, Meyer de
Schauensee 1970), and this is reflected in the linear sequence of the AOU
(1998). Genetic data (Griffiths et al.
2007, Raposo do Amaral et al. 2009) also indicate that it is not closely
related to any buteonine genera, where traditionally place, but rather closer
to kites. SACC proposal
needed.
Olson
(1982) used toe morphology to predict that Busarellus
is not closely related to buteonine hawks but rather to Old World kites. Raposo do Amaral et al.’s (2009) phylogeny
(see proposals 459, 2460) clearly indicated that Busarellus is not within the buteonines;
because their analysis targeted that group, only a few outgroup taxa were
used. Nevertheless, a well-supported
node group Busarellus with Geranospiza and Rostrhamus sociabilis to
the exclusion of all the buteonines as well as Ictinia, Old World Butastur;
and Haliaeetus; sampling did not
include the taxa mentioned by Olson, but regardless, Busarellus cannot be considered buteonine. The relevant portion of their tree (from
their Fig. 3) is pasted in here:
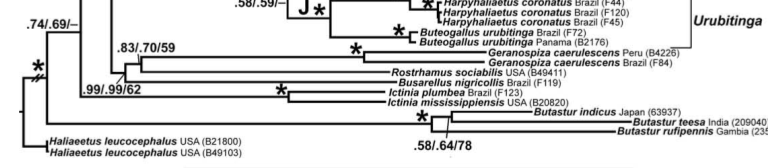
Therefore,
at present, there is no evidence that Busarellus
is a buteonine hawk. Although which
non-buteonines are actually its closest relatives cannot be determined without
additional taxon sampling, it is clear that it must be moved elsewhere. Griffiths et al. (2007) sampled extensively outside
the buteonines but sequenced only one (nuclear) gene (RAG-1). Their maximum-likelihood tree places Busarellus with Butastur and Ictinia;
however, support for the structure of this whole region of the tree is weak to
nonexistent, and that further sampling could show it to be the sister of any of
the genera in the tree from Rostrhamus
to Geranospiza, and possibly even
others more basal in the group:
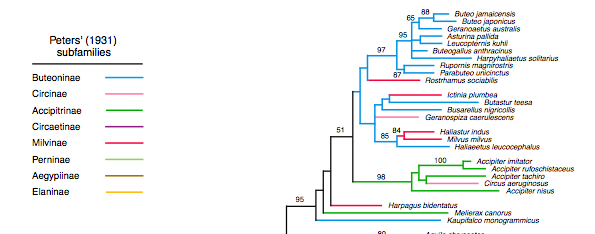
Thus,
we could place it anywhere in the current sequence of non-elanine kite genera:
Rostrhamus sociabilis Snail Kite
Helicolestes hamatus Slender-billed Kite
Harpagus bidentatus Double-toothed Kite
Harpagus diodon Rufous-thighed Kite
Ictinia mississippiensis Mississippi Kite
Ictinia plumbea Plumbeous Kite
Note
that the RAG-1 tree conflicts strongly with the Raposo do Amaral et al. tree in
terms of which genera are closest to Busarellus:
Rostrhamus or Geranospiza but definitely not Ictinia
in Raposo do Amaral et al., but most likely (statistically) Ictinia or Geranospiza and much less likely Rostrhamus in Griffiths et al.
Which one is correct requires additional taxon and gene sampling. If part b (below) is rejected, arbitrarily
placing it after Ictinia in our
current sequence in hopes that the unsupported structure in RAG-1 tree turns
out to be correct would be my recommendation.
We
could also place it next to Geranospiza,
currently between Accipiter and Leucopternis, but that placement of Geranospiza makes little sense in light
of the results of Griffith et al. (2007).
This leads to the second part of the proposal: an overhaul of the
sequence of genera
Part b: rearrange sequence of genera to
follow Griffiths et al. (2007)
Here
I propose that we change our sequence so that it corresponds more closely with
that of Griffiths et al. (2007), which we already used to move some of the kite
genera (e.g., Elanus and Gampsonyx) to the beginning of the
sequence. My rationale is that (a) as
long as Busarellus must be moved, and
(b) the likely close relatives to Busarellus
are separated by monophyletic groups such as Accipiter + Circus, we
might as well adopt the general structure of the Griffiths et al. tree. This would also place Geranospiza among the other candidate genera for the sisters to Busarellus (instead of between Accipiter and the buteonines). There will always be questions about trees
based on a single gene, but the support for the deep nodes in the RAG-1 tree
are strong, and the groupings themselves are all sensible. Additional data might require changes to the
sequence, but basing our sequence on the RAG-1 tree reflects the best data
currently available. Finally, the
current linear sequence of groups of genera is based on mostly historical
momentum, which is largely a concoction of antiquated ideas about “primitive”
and “advanced” hawks – therefore, ANY new sequence based on modern data would
have more published support and testable hypotheses than the historical one.
The
RAG-1 tree:
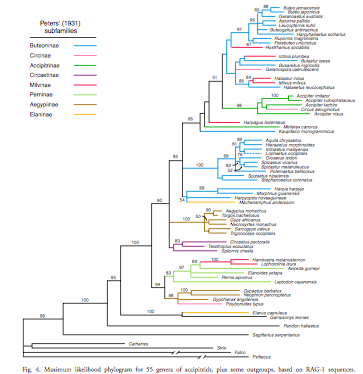
and
might be better visualized from their more schematic version:
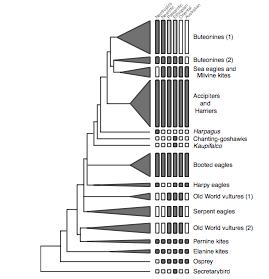
Adopting
the Griffiths et al. tree would involve one major rearrangement: bringing
forward in the sequence the eagle genera Morphnus,
Harpia, and Spizaetus, thus leaving the buteonines last (see proposals 459 and 460 for the proposed realignments in those
genera). The only other “major” change
would be the above-mentioned move of Geranospiza
forward to the kites.
Our
current sequence is as follows (with Busarellus
and putative relatives in red):
Elanus
Gampsonyx
Chondrohierax
Leptodon
Elanoides
Rostrhamus
Helicolestes
Harpagus
Ictinia
Circus
Accipiter
Geranospiza
Leucopternis
Buteogallus
Harpyhaliaetus
Busarellus
Geranoaetus
Parabuteo
Buteo
Morphnus
Harpia
Spizaetus
A sequence consistent
with Griffiths et al. that minimizes the number of changes is (note that
proposals 459 and 460 would affect the buteonine genera Leucopternis
through Buteo, but the group itself would stay intact); taxa whose
position is changed are shown in blue:
Elanus
Gampsonyx
Chondrohierax
Leptodon
Elanoides
Morphnus
Harpia
Harpagus
Circus
Accipiter
Rostrhamus
Helicolestes
Busarellus
Ictinia
Geranospiza
Leucopternis
Buteogallus
Harpyhaliaetus
Geranoaetus
Parabuteo
Buteo
Thus, the beginning and
end of the sequence stays relatively stable.
The harpy eagles and booted eagles are brought forward, as is Harpagus
slightly and Circus + Accipiter slightly. The milvine kites, which would include Rostrhamus
through Geranospiza, are all now together, although whether they form a
monophyletic group is questionable from the RAG-1 tree. The main accomplishment is bringing Busarellus
out of the buteonines, placing it among its potential relatives. The proposed new sequence also rescues the
harpy eagle types from any association with the monophyletic buteonines. I considered moving Harpagus and Rostrhamus
(with unproven close relative Helicolestes) slightly to match the RAG-1
branching pattern, but there is little or no support for the nodes on that
section of the tree. Tweaks welcomed.
Recommendation: The proposed sequence is consistent
with the phylogenetic hypotheses of Griffiths et al.’s RAG-1 tree, and
extricates Busarellus from the
buteonines. Pending tweaks, I recommend
a YES vote to incorporate the best phylogenetic data available into our
sequence. The vote is broken into two
parts:
a)
= to move Busarellus out of buteonines
b) = to use proposed sequence above.
Literature Cited:
GRIFFITHS, C. S., G.
F. BARROWCLOUGH, J. G. GROTH, AND L. MERTZ. 2007. Phylogeny, diversity and
classification of the Accipitridae based on DNA sequences of the RAG-1 exon. J.
Avian Biology 38: 587-602.
OLSON, S. L. 1982. The
distribution of fused phalanges of the inner toe in the Accipitridae. Bull.
British Ornithol. Club 102: 8-12.
RAPOSO DO AMARAL, F.,
F. H. SHELDON, A. GAMAUF, E. HARING, M. RIESING, L. F. SILVEIRA, AND A.
WAJNTAL. 2009. Patterns and processes of diversification in
a widespread and ecologically diverse avian group, the buteonine hawks (Aves,
Accipitridae). Molecular Phylogenetics and Evolution 53: 703-715.
Van
Remsen, August 2010
_____________________________________________________________________________
Comments
solicited from Fabio Raposo: “Two important references may make the
difference here. Based on a representative taxonomic sampling of the
family Accipitridae, Lerner & Mindell 2005 recovered species of
Haliaeetus, Ichthyophaga, Haliastur and Milvus as
the closest relatives of the "core" buteonine hawks + Ictinia,
Geranospiza and Rostrhamus (Fig. 1 of that paper), based on two
mitochondrial markers. A smaller taxonomic sampling of the
mitochondrial dataset plus one nuclear intron provides a similar
picture with even better support (Fig. 2). "Kites" seem to represent
an artificial group, since kite genera appear all over their accipitrid tree. So,
it is possible that Geranospiza, Rostrhamus, Ictinia and Butastur
are indeed better thought as buteonine hawks - and Busarellus
and Butastur also seem to be part of this clade (see Lerner et
al. 2008, besides Amaral et al. 2009).
Lerner, H.R., Mindell, D.P., 2005. Phylogeny
of eagles, Old World vultures, and other Accipitridae based on nuclear and
mitochondrial DNA. Molecular Phylogenetics and Evolution 37, 327–346.
Lerner,
H.R.L., Klaver, M.C., Mindell, D.P., 2008. Molecular phylogenetics of the
buteonine birds of prey (Aves, Accipitridae). Auk 125, 304–315.