Proposal (466) to South American Classification Committee
Transfer Caprimulgus rufus and Caprimulgus sericocaudatus
to the genus Antrostomus
Han et al. (2010) published a molecular based
phylogeny of Caprimulgidae from more than 60% of caprimulgid species and 14 of
16 currently recognized genera. The taxon sampling was broad and included all
morphologically divergent lineages. New World taxa were particularly well
represented. DNA sequences were collected from the entire mitochondrial DNA
cytochrome b gene and parts of two nuclear genes (myelocytomatosis viral
oncogene homolog and growth hormone). All analyses of the 72 ingroup plus
outgroup taxa were conducted on the 4179 characters. The Han et al. (2010)
phylogeny was well resolved and in substantial agreement with all previous
molecular work on the family (Sibley and Ahlquist, 1990; Mariaux and Braun,
1996; Barrowclough et al., 2006; Larsen et al., 2007; Braun and Huddleston,
2009).
In proposing a new classification, Han
et al. (2010) used the following criteria: foremost, all named taxa represented
monophyletic groups. Second, for stability of the named taxa, whenever
possible, currently recognized taxa were retained, and when more than one
partitioning scheme for a clade was plausible, they opted for the one that was
more likely to remain viable in the face of new data.
Within core caprimulgids, four strongly
supported major clades (3 New World, 1 Old World) provided a natural partitioning
scheme, but numerous taxa needed to be reassigned to reflect the non-monophyly
of the current genus Caprimulgus. Additionally, a number of small or
monotypic genera should be subsumed. Caprimulgus Linnaeus 1758 should be
restricted to the Old World clade, since the type specimen for the genus is Caprimulgus
europaeus.
Here is the tree from Han et al.:
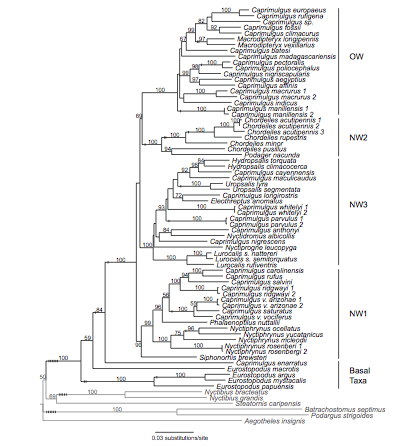
Proposed
change:
Of taxa that are germane to SACC within Han et
al.’s (2010) New World clade 1, Nyctiphrynus remains as currently
constituted. Caprimulgus rufus is transferred to the genus Antrostomus
Bonaparte 1838, which has priority among available names for this clade (Peters
1940). Although not included in the Han et al. (2010) study, Caprimulgus
sericocaudatus is also assigned to Antrostomus. The allocation of sericocaudatus
to Antrostomus is based on its placement in a species group with salvini
and badius by Cleere (1999), and more importantly, an unpublished
molecular data set using different genes than Han et al. (2010) by Snorri
Sigurdsson and Joel Cracraft (2010) demonstrated that C. sericocaudatus
is sister to C. rufus and C. carolinensis and thus belongs in the
Antrostomus clade. Although C.
carolinensis is a North American breeding species occurring in SACC area
only in winter, it is the sister species to C. rufus and should thus
also follow that species into Antrostomus.
References:
Barrowclough,
G.F., Groth, J.G., Mertz, L.A. 2006. The RAG-1 exon in the avian order
Caprimulgiformes: phylogeny, heterozygosity, and base composition. Mol.
Phylogenet. Evol. 41:238–248.
Braun,
M.J., Huddleston, C.J. 2009. A molecular phylogenetic survey of caprimulgiform
nightbirds illustrates the utility of non-coding sequences. Mol. Phylogenet.
Evol. 53:948–960.
Cleere,
N. 1999. Family Caprimulgidae (Nightjars). In: del Hoyo, J., Elliott, A.,
Sargatal, J. (Eds.), Handbook of the Birds of the World, Barn Owls to
Hummingbirds, vol. 5. Lynx Edicions, Barcelona, pp. 302–386.
Han,
K.-L., Robbins, M.B., Braun, M.J. 2010. A multi-gene estimate of phylogeny in
the nightjars and nighthawks (Caprimulgidae). Mol. Phylogenet. Evol.
55:443-453.
Larsen,
C., Speed, M., Harvey, N., Noyes, H.A. 2007. A molecular phylogeny of the
nightjars (Aves: Caprimulgidae) suggests extensive conservation of primitive
morphological traits across multiple lineages. Mol. Phylogenet. Evol.
42:789–796.
Mariaux,
J., Braun, M.J. 1996. A molecular phylogenetic survey of the nightjars and
allies (Caprimulgiformes) with special emphasis on the potoos (Nyctibiidae).
Mol. Phylogenet. Evol. 6:228–244.
Peters,
J.L. 1940. Checklist of Birds of the World. Harvard University Press,
Cambridge.
Sibley,
C.G., Ahlquist, J.E. 1990. Phylogeny and Classification of Birds: A Study in
Molecular Evolution. Yale University Press, New Haven, London.
Sigurdsson, S., Cracraft, J. 2010. A species-level phylogeny of
the nightjars (Caprimulgidae). Poster at
International Ornithological Congress, Campos de Jordão, Brazil.
Mark Robbins, September 2010
___________________________________________________________________________
Comments from Stiles: “YES
– this clade also seems strongly supported, with only Siphonorhis and Nyctiphrynus perhaps
meriting generic separation from Antrostomus
(I note that carolinensis, not
mentioned in the proposal, also falls into Antrostomus).”
Comments from Nores: “YES. Es evidente en el
análisis molecular de Han et al. (2010) que C. rufus y C. carolinensis forman
un clado que incluiría también a C.
sericocaudatus (en base a un análisis no publicado de Snorri
Sigurdsson and Joel Cracraft). Pero ver respuesta a propuesta #441.”
Comments from Remsen: “YES. There really aren’t any other options, although Mark did not address what will happen in terms of classification with the whip-poor-will group, presumably because it is extralimital. Given the overall similarity in voice and morphology, I would favor expanding the boundaries of this genus to include the node with support value 96 that includes not only the whip-poor-will group but also Phalaenoptilus. To maintain the latter as a monotypic genus when in the previous proposal Lurocalis and, for example “C.” parvulus are merged into one genus is inconsistent in my opinion. Even including Nyctiphrynus with Antrostomus would produce a less heterogeneous group than broad Hydropsalis.”
Comments from Zimmer: “YES.
This move seems strongly supported by the molecular data and makes sense
on the grounds of vocal and morphological characters. I would echo Van’s suggestion to include the
extralimital whip-poor-will group (and include Phalaenoptilus) in Antrostomus,
although I wouldn’t go so far as to include Nyctiphrynus,
which, to me, seem to be pretty different in a number of respects.”
Comments from Pacheco: “YES. Considero a proposta bem apoiada pelos dados disponíveis.”
Comments from Jaramillo: “YES. The molecular data are strong, and vocalizations and other natural history considerations suggest that this is a solid group, and not part of the true Caprimulgus of the Old World. I also question why the whip-poor-will group was left out of the discussion, presumably as they are not Neotropical species, but it would be good to know that it isn’t for a taxonomic reason we should be aware of.”
Comments from Pérez: “YES,
I think molecular data presented by Han et al. (2010) strongly support this
group. I like Van’s proposal but also expanding Antrostomus would need to consider Phalaenoptilus within this genus, a decision to which I don’t have
a strong opinion. Phylogenetically and
based on vocal (based on Kevin’s comments) and morphological grounds, it would
make sense but considerations of its physiology might support placement in its
own genus. Either one would make sense depending on the weight we give to each
character and on our definition of a genus (which could be based on different
criteria). Maybe we need to discuss these issues to be consistent when
evaluating different proposals. Going back to Van’s proposal, excluding Phalaenoptilus from Antrostomus would force to define another genus for the node
including C. vociferus
(whip-poor-will) as there is no strong support for the node suggesting both
groups of ex-Caprimulgus are sister
taxa.”