Proposal (534) to South American
Classification Committee
Changes
to Pipridae genera and sequence
A. Changes to generic allocation
Background: The phylogeny of the manakins has been
investigated substantially over the last 20 years or so, and the arrangement we
follow does not completely correspond to the results of either morphological or
molecular studies done on the family.
Recent molecular studies suggest that we need to make at least some
changes in the allocation of species to genera.
Based on
syringeal morphology, Prum (1992) concluded that the large manakin genus Pipra was polyphyletic and split the
small manakins of the serena group
out into the genus Lepidothrix, and
recognized Dixiphia for the
White-crowned Manakin, P. pipra. The recognition of Lepidothrix has been accepted by nearly all subsequent authors and
was part of the original base taxonomy for SACC based on the treatment in
Dickinson (2003). Dixiphia has not been accepted by SACC including a recent proposal
that considered both Prum’s syringeal data and molecular data from Rego et al
(2007). Since that time, two additional
molecular studies (Tello et al 2009) and McKay et al (2010) provide additional
data regarding relationships in Pipridae.
New information: Since 2007,
three studies using DNA sequences have provided data and analysis that bears on
this issue. Rego et al (2007) used
mitochondrial cytochrome B and rRNA 16S to examine relationships within the
Pipridae. They sampled 18 species
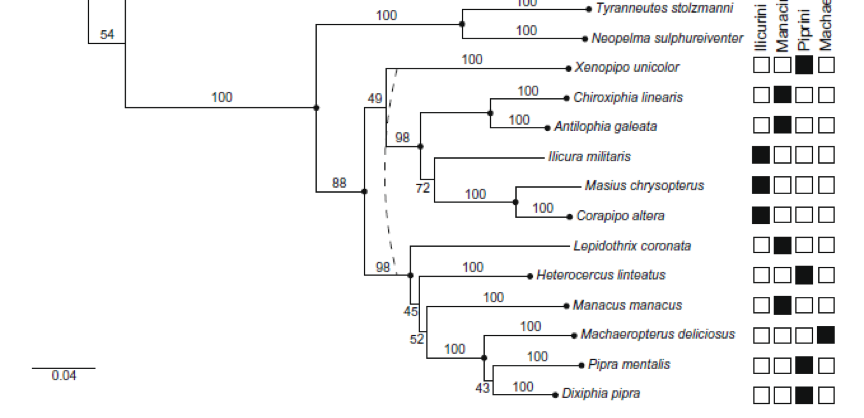
representing 13 genera. McKay et al.
(2010) used two mitochondrial genes (nd2 and col) and a nuclear intron Musk
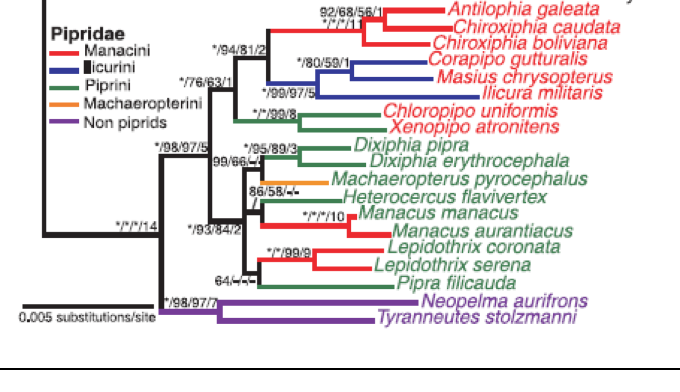
intro 3 to look at Pipridae, sampling 14 species representing 14 genera. Tello et al. (2009) used two nuclear genes
(RAG-1 and RAG-2) to look at the broader radiation (Tyrannides) from Tyrannidae
though Cotingidae to Pipridae. They sampled a total of 19 manakin species
including representatives of all of the relevant genera.
The relevant
portions of the trees for Pipridae from all three molecular studies are
reproduced below. One thing you will
note is that the taxon sampling is not close to complete in any of these
studies, but that there is a fair amount of complementarity among the taxa used
in the studies. There are a number of
things going on, and certainly not complete agreement among the studies. All three studies identify a clade of what I
would call classic manakins (plus the weird Heterocercus),
including the genera Pipra, “Dixiphia”,
Heterocercus, Manacus, Lepidothrix, and Machaeropterus. There is some disagreement on the
relationships among these genera.
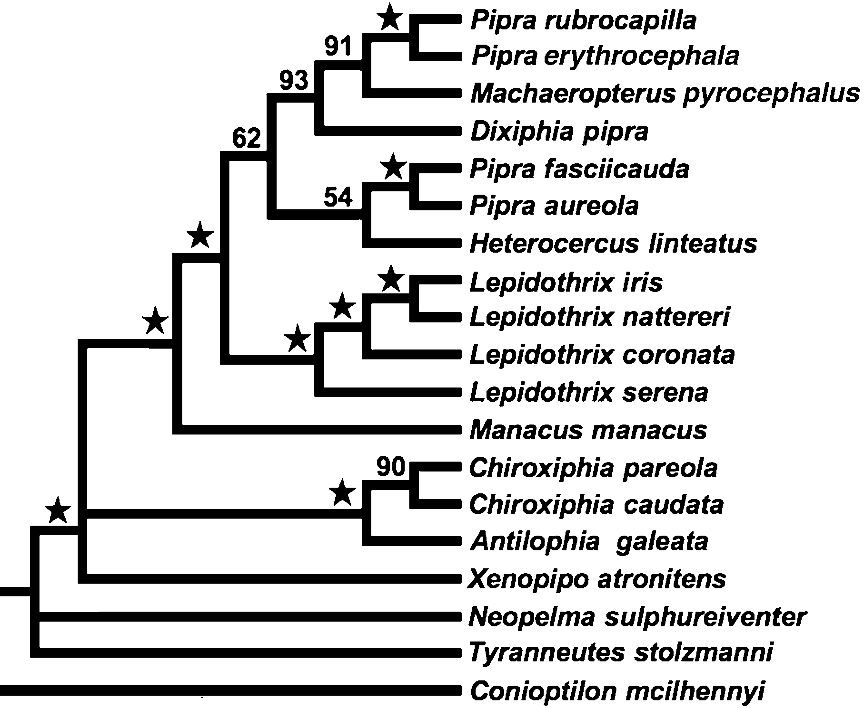
However, one subclade is consistently returned by all 3 studies. That clade contains Machaeropterus, Pipra (or
Dixiphia) pipra, and the cornuta species
group of Pipra (represented by rubrocapilla, erythrocephala and/or mentalis in the three studies). The remaining species of Pipra (the aureola species group) do not cluster with these taxa in the Rego
et al (2007) or the Tello et al (2009) studies.
Unfortunately, McKay et al. (2010) lacked a representative of the aureola group.
The clade with Machaeropterus, Pipra pipra and the cornuta
group does not have a consistent topology in the 3 studies. Rego et al. (2007) have Machaeropterus interposed between Pipra pipra and the cornuta
group, whereas the other two studies have Machaeropterus
at the base of the clade.
It
appears from these studies that the genus Pipra
even with the removal of Lepidothrix and
“Dixiphia” pipra is not monophyletic.
The generic name Pipra belongs
with the aureola group. That group seems to form straightforward
clade. There are 3 generic names associated with the clade that includes the cornuta group currently placed in Pipra.
Dixiphia is the oldest name,
and P. pipra is the type. Machaeropterus
applies to those species currently in that genus and Ceratopipra would be the appropriate name for the cornuta group.
There
would seem to be 3 possible treatments for the Dixiphia, Machaeropterus, and Ceratopipra
clade. A) The taxa could all be placed
in the genus Dixiphia, the oldest
name for the group. B) The three names
available could all be used, recognizing Machaeropterus
as currently defined, Dixiphia as
monotypic, consisting of just pipra,
and Ceratopipra for the cornuta group. C) Retain Machaeropterus
and place pipra and the cornuta group in Dixiphia. The first two
treatments are consistent with all three molecular studies. The third, however, conflicts with the tree
of Rego et al. Thus, I think the two
appropriate options are using Dixiphia for
all these taxa or the 3-genera treatment.
Because Machaeropterus stands
out morphologically, in plumage pattern and behavior from the other species, I
am disinclined to place all species in Dixiphia.
Rego et al. (2007) tree:
Tree from Tello et al (2009):
Tree from McKay et al (2010):
Recommendation:
The
molecular data clearly indicate that Pipra
as currently constituted is not monophyletic.
So I recommend removing Pipra
pipra, cornuta, chloromeros, rubrocapilla, and erythrocephala from Pipra. Two possible treatments for the clade found in
all three studies including these species plus the genus Machaeropterus are possible:
A) All taxa placed in the genus Dixiphia,
the oldest name for the group. B) The
three names available could all be used, recognizing Machaeropterus as currently defined, Dixiphia as monotypic, consisting of just pipra, and Ceratopipra for
the cornuta group. Because Machaeropterus
stands out morphologically, in plumage pattern and behavior from the other
species, I am disinclined to place all species in Dixiphia, and recommend choice B.
B. Sequence of genera in Pipridae
Assuming that
SACC splits Pipra, and basically
accepts the results of these molecular studies, we need to adjust the order of
genera in Pipridae to reflect the results from these molecular studies. All 3 studies have Tyranneutes and Neopelma
basal. They also identify two clades,
one consisting of Ilicura, Masius,
Corapipo, Antilophia, Chiroxiphia, and Xenopipo. The other clade contains the remaining
genera: Pipra, Lepidothrix, Manacus,
Heterocercus and Dixiphia (sensu
lato). The topology of the first clade
seems well-established. Xenopipo is basal, Antilophia and Chiroxiphia
are sisters, Masius and Corapipo are sisters, and Ilicura is sister to them.
The other clade
has much more variation across the 3 studies, and bootstrap values for much of
the structure are poor. However, as
noted in part A, a clade containing Machaeropterus,
Dixiphia, and Ceratopipra is returned by all three studies, and a sequence of
those taxa with Dixiphia between the
other two is consistent with all of the topologies. The remaining genera, Pipra, Lepidothrix, Manacus, and
Heterocercus, have very different arrangements in the three studies. A sequence of Manacus, Heterocercus, Pipra, and Lepidothrix seems like it would best reflect the potential
relationships suggested in these studies.
So I recommend
we place the genera of Pipridae in the following sequence, which is consistent
with the molecular topologies and maintains the current sequences as much as
possible:
Neopelma
Tyranneutes
Ilicura
Masius
Corapipo
Antilophia
Chiroxiphia
Xenopipo
Machaeropterus
Dixiphia
Ceratopipra
Manacus
Heterocercus
Pipra
Lepidothrix
References:
Dickinson,
E. C. (ed.). 2003. The Howard and Moore complete checklist of
the birds of the World, Revised and enlarged 3rd Edition. Christopher Helm,
London, 1040 pp.
McKay, B. D., F. K. Barker, H. L.
Mays, Jr., S. M. Doucet, and G. E. Hill.
2010. A molecular phylogenetic
hypothesis for the manakins (Aves: Pipridae). Molecular Phylogenetics and
Evolution 55:733-737.
Prum, R. O. 1992. Syringeal morphology, phylogeny, and
evolution of the Neotropical manakins (Aves: Pipridae). American Museum Novitates 3043.
Rego, P. S., J. Araripe, M. L. V.
Marceliano, I. Sampaio, and H. Schneider.
2007. Phylogenetic analyses of
the genera Pipra, Lepidothrix and Dixiphia (Pipridae, Passeriformes) using partial cytochrome b and
16S mtDNA genes. Zoologica Scripta
2007:1-11.
Tello, J. G., R. G. Moyle, D. J.
Marchese, and J. Cracraft. 2009. Phylogeny and phylogenetic classification of
tyrant-flycatchers, cotingas, manakins and their allies (Aves: Tyrannides). Cladistics 25:429-465.
Doug Stotz, July 2012
_________________________________________________________________________________________________________________
Comments from Zimmer:
“A. YES. I would concur with
Doug’s rationale for choice “B”: That
is, to retain Machaeropterus as
currently defined, restrict Dixiphia
to pipra (which is more than one
species anyway), and move the cornuta group
(including chloromeros, in addition
to the species sampled) to Ceratopipra. Given the distinctiveness of Machaeropterus, I just can’t see lumping
everything into Dixiphia. B. YES
to Doug’s proposed sequence, which makes the most sense while minimizing
upheaval.”
Comments
from Stiles: “A. YES to B, especially as
the display behavior of pipra is
really quite different from that of at least erythrocephala and mentalis
and setting it apart also fits Prum’s morphological data.”
Comments from Pacheco: “A. Yes. B. YES. A sequência me parece
consistente com os dados recentes disponíveis, incluindo uma proximidade na
sequência entre Dixiphia e Machaeropterus.”
Comments from
Stiles: “YES,
including the recognition of three genera: Ceratopipra,
Dixiphia and Machaeropterus.”
Comments from Pérez-Emán: “A: YES. Data are consistent with recognition of
Dixiphia, Machaeropterus, and Ceratopipra.
B: YES. Sequence is consistent with what
we know so far about these taxa, while creating the fewest changes.”
Comments from Jaramillo: “YES - for both A and B. The final arrangement
makes sense and is based on various solid data.”
Comments from Nores: “Changes to generic allocation. YES, choice B. The
resurrection of the genus Ceratopipra is consistent with the Rego et
al. study, but conflicts with the trees of Tello et al. and the McKay et al.
Note, however, that this does not mean eliminating
the genus Machaeropterus,
which groups a set of species quite similar in color
to each other and different from the rest.
Sequence of genera in Pipridae. NO. Following the criteria of the taxon that splits first
(presenting the lesser number of ancestors, that is, internal nodes) being
placed at the top of the sequence and so on, we obtain the following sequences
of the trees of Rego et al., Tello et al. McKay et al. and a new sequence.
SACC Stotz Rego et al. Tello et al. McKay et al.
Neopelma Neopelma Neopelma Neopelma Neopelma
Tyranneutes Tyranneutes Tyranneutes Tyranneutes Tyranneutes
Ilicura Ilicura Xenopipo Xenopipo Lepidothrix
Masius Masius Antilophia Antilophia Heterocercus
Corapipo Corapipo Chiroxiphia Chiroxiphia Manacus
Machaeropterus
Antilophia Manacus Ilicura Machaeropterus
Lepidothrix Chiroxiphia Lepidothrix Corapipo Dixiphia
Manacus Xenopipo Heterocercus Masius Ceratopipra
Antilophia
Machaeropterus Pipra Heterocercus Xenopipo
Chiroxiphia Dixiphia Dixiphia Manacus Antilophia
Xenopipo Ceratopipra Machaeropterus Lepidothrix Chiroxiphia
Heterocercus Manacus Ceratopipra Machaeropterus Ilicura
Pipra Heterocercus
Dixiphia Corapipo
Pipra
Ceratopipra Masius
Lepidothrix
Proposed new sequence:
Neopelma
Tyranneutes
Xenopipo
Antilophia
Chiroxiphia
Ilicura
Corapipo
Masius
Manacus
Heterocercus
Pipra
Lepidothrix
Machaeropterus
Dixiphia
Ceratopipra