Proposal (537) to South American Classification Committee
Elevate Pauxi unicornis koepckeae to species status
Effect of Proposal:
If it passes, this proposal would result in treatment of Pauxi unicornis
koepckeae as a separate species from P. u. unicornis (Horned
Curassow).
Background: Current classifications
typically treat Pauxi unicornis as consisting of two subspecies, with
disjunct ranges: nominate unicornis in the Andean foothills of Bolivia,
and koepckeae, known only from the isolated Sira range in Peru, ~1000 km
north (and ~60 km east from the nearest Andean slopes over 1000 m, Terborgh
& Weske 1971). The two
subspecies are differ in morphology (Weske & Terborgh 1971, Gastañaga et al. 2011), vocalizations, and behavior (Gastañaga et al. 2011).
New information:
Gastañaga et al. (2011) studied vocal, behavioral, ecological, and morphological
differences between the two populations of P. unicornis, as follows:
Vocal differences – koepckeae has a low booming vocalization that lacks the final emphatic far
carrying note that is the most distinctive feature of the song of unicornis. The booming sequence of koepckeae
is a series of up to four notes followed by a pause of approximately 2 sec,
repeated every 4-5 sec. In
contrast the unicornis booming
sequence comprises four phrases totaling eight notes that lasts 8-10 sec and is
repeated every 15 sec, and also has a very distinctive and very loud final
sharp ‘BMM!’ note that koepckeae
lacks (Figure and Table below). These vocal differences are based on N = 97
encounters with singing individuals (N = 25 koepckeae
and N = 72 unicornis) between 1999
and 2008.
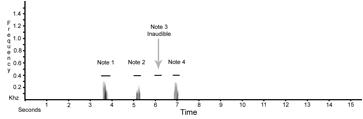
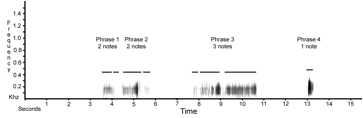
Sonograms of koepckeae (upper)
and unicornis (lower). The song of koepckeae has a phrase of four notes
(indicated by the black horizontal lines) with the first note somewhat louder
than the following 3 notes. In the
sonogram the third note was so quiet that it could not be separated from the
background noise of the recording although it was audible to the observers in
the field. The song of unicornis has eight notes (indicated by
the black horizontal lines), forming the four phrases shown in the
sonogram. The song of unicornis is three times longer, with
the final distinctive and far-carrying note much louder and this is completely
absent from the song of koepckeae.
Variation
in vocal signatures of P. u. koepckeae and P. u. unicornis.
|
Pauxi form |
Final loud
note |
Phrase duration |
Pause duration |
Number of phrases |
Number of notes |
Tail
gesture during
alarm calls |
N |
|
koepckeae |
No |
3 sec |
2 sec |
1 |
4 |
horizontal
fanning |
25 |
|
unicornis |
Yes |
8-10 sec |
5-7 sec |
4 |
8 |
vertical
pumping |
72 |
The song of the only other
curassow found in the Sira Mountains, Mitu
tuberosum, has a similar number of notes and length of song to unicornis, and its song is much more
similar to unicornis than koepckeae is to unicornis.
The ‘Ksop!’ alarm call was accompanied by a horizontal
fanning of the tail feathers concordant with each call in koepckeae; unicornis
accompanies each alarm call by pumping the tail up and down concordant with
each call.
Habitat and altitudinal differences – koepckeae is resident in cloud forest (1100-1600 m), whereas unicornis occurs in humid forest on the edge of tropical lowlands
and lower montane forest (400-1100 m).
The Sira Mountains are
isolated from the Andes, characterized by habitat zones that are at lower
altitudes than the Andes; the cloud forest to montane forest transition is at
1050 m, which is several hundred meters lower than this transition at
comparable locations in the main Andes (low of 1380 m, Terborgh & Weske 1975).
Therefore the difference in habitats occupied by the two taxa is even
greater than is suggested by a simple comparison of altitudinal ranges.
Morphological differences – As noted by (Weske & Terborgh 1971) there are striking differences
in casque form, with a flattened shield-like ellipsoid in koepckeae and an erect cone-shaped horn in unicornis (see Figure below), as well as koepckeae having less white at the tip of the tail than either unicornis or pauxi. We have also confirmed the casque shape
of koepckeae (N = 14 observations)
differs consistently from unicornis
(N > 30 observations) in the field, with all observation recording the
upright horn in unicornis and the
flat shield in koepckeae and no
intermediate casque forms recorded in either population.



P. koepckeae (Peru) P. unicornis (Bolivia)
Although
Weske & Terborgh (1971) offer measurements of casque diameter and culmen tip to top of casque for koepckeae, they were not compared to
other congeners, which we offer in the Table below. We note non-overlapping measurements between koepckeae and its two congeners for
culmen tip to top of casque, maximum
diameter of casque, and casque length. For culmen tip to top of
casque and casque length the differences between koepckeae and unicornis
are greater than the differences between pauxi
and unicornis. We also note the
measurements are not consistent with a latitudinal cline across the 3 species
as the koepckeae measurements are not
intermediate between those of unicornis
in Bolivia and pauxi in
Colombia/Venezuela.
Variation
in casque morphology among separate forms of Pauxi.
|
Pauxi |
Culmen
tip to top of casque |
Max
diameter of
casque |
Casque
length |
|
koepckeae |
84.2 (83.6-84.9) |
21.4 (20.4-22.5) |
45.3 (45.0-45.7) |
|
unicornis |
99.3 (98.9-99.7) |
24.6 (23.0-26.3) |
56.8 (55.0-58.6) |
|
pauxi |
95.2 (87.0-104.1) |
36.3 (29.8-44.0) |
60.5 (57.2-62.3) |
Discussion: As noted above, the
vocalizations of koepckeae differ
strongly from those of unicornis, and
in many ways the vocalizations of unicornis
are more similar to those of Mitu
tuberosum than koepckeae. For ranking allopatric taxa under the
Biological Species Concept, differences of this magnitude in voice are
typically considered strong evidence for ranking the taxa as species. The differences in casque size and
shape might also be associated with divergence to the point of being a barrier
to gene flow if these structures are associated with mate selection. Although differences in habitat and
elevation are not species-level characters per se, they are at least consistent
with species-level differences.
Recommendation: A “YES” vote. Although
assessment of species limits among allopatric taxa is problematic, we think
that the degree of differentiation between these widely separated populations
is consistent with the level of differences associated with taxa ranked as
species in the Cracidae. For
example, these differences are arguably greater than those between taxa treated
as separate species in Penelope, Aburria/Pipile, and Ortalis.
English names: We recommend “Sira Curassow” for koepckeae
and retaining “Horned Curassow” for unicornis. Although there are advantages to
creating two new names for the split species, we prefer the simpler names to
something like Northern Horned Curassow and Southern Horned Curassow.
References:
Gastañaga, M. 2006. Peruvian Horned Curassow (Pauxi unicornis koepckeae) rediscovered in the Sira Mountains, Peru.
Bulletin of the Cracid Specialist Group 22: 10-23.
Gastañaga, M., A. B. Hennessey, & R. MacLeod. 2007. Rediscovery of
Southern Horned Curassow Pauxi unicornis
koepckeae in Cerros del Sira, Peru. Cotinga 28: 63-66.
Gastañaga-C., M., R. MacLeod, D.M. Brooks and B. Hennessey. 2011. Distinctive morphology, ecology, and first vocal descriptions of
Sira Curassow (Pauxi [unicornis] koepckeae): evidence for
species rank. Ornitol. Neotrop. 22: 267-279.
Graham, J.G. 2009. A new specimen of Southern Horned Curassow Pauxi unicornis from Peru. Cotinga 31:
73.
Gúzman, E., D. M. Brooks, & G. Sedaghatkish. 1999. Notas sobre la
historia natural de los crácidos albergados en el Museo “Noel Kempff Mercado”,
Santa Cruz, Boliva, con notas sobre la taxonomía de las pavas del genero Pipile. Bulletin of the Cracid
Specialist Group 8: 20-28.
Herzog, S. K., & M. Kessler. 1998. In search of the last
Horned Curassows Pauxi unicornis in
Bolivia. Cotinga 10: 46-48.
Maillard-Z.,
O. 2006. Reciente espécimen de la Pava Copete de Piedra (Pauxi unicornis) para Bolivia. Kempffiana
2: 95-98.
Weske, J.S. and J.W. Terborgh. 1971. A new subspecies of curassow of the
genus Pauxi from Peru. Auk 88:
233-238.
Daniel Brooks, Ross MacLeod, and Melvin Gastañaga-C.
August 2012
Comments
from Remsen: “YES. I think the proposal clearly places burden-of-proof on
treatment of koepckeae at the subspecies rank. In fact, I think that species rank is better-justified than
it is for a number of allopatric species in the Cracidae.”
Comments from Jacob
Socolar: “I want to offer three brief comments. I do not expect any of
these comments to substantially influence the committee members' decisions, but
perhaps it could somehow be useful to mention these points:
“--On
August 7, 2010, I observed a Pauxi [unicornis] koepckeae at great
length at 1650 masl on Terborgh and Weske's original transect of the Sira.
This individual clearly had white tipping to all retrices, and I do not
believe that the extent of white in the tail is a consistent difference between
koepckeae and nominate unicornis.
“--The
casque morphology is somewhat variable in koepckeae. In
particular, I have in my possession a very poor photo of a P. [u.] koepckeae (which
I can send to anyone who wants to see it) that shows an individual of koepckeae with
a casque that appears to approach Pauxi pauxi in shape. The
casque morphology in Pauxi pauxi is also variable. Among the
specimens at the Smithsonian, the only gender/subspecies combination with multiple
specimens involves three female P. p. gilliardi, and their
casques are tremendously variable in size and shape, apparently due to damage
and wear during the birds' lifetimes.
“--Whether
koepckeae is truly sister to P. unicornis or P.
pauxi is not entirely clear to me.
“While
I think that additional data (likely including genetic work and also a further
attempt to follow up on Ted Parker's possible record) are required to
thoroughly understand the relationships between the various taxa in the genus Pauxi,
I agree with the recommendation for a 'yes' vote on this proposal.”
Additional comments
from Brooks and MacLeod: “Jacob, thanks for
your comments, here are some brief responses:
“The individual you saw at Terborgh and Weske's site was probably in
molt and simply shed the central rectrix. Mel and Ross found two shed
rectrices in the Sira in mid July 2007, which is close to your date of 7
August. The Holotype at AMNH deposited by Weske and Terborgh from the
same site where you saw the bird in question also lacks a white tip centrally.
- “We're
not surprised about the casque variation in the small series of female P. p.
gilliardi, as variation increases with size of a given character, and P. pauxi has a large casque. More
importantly there is some ontogenetic variation in casque size of different age
classes since Pauxi reaches adult
body size within a year, but the casque doesn't attain full size for 2-3 years.
Despite the potential for size variation due to growth we've seen at
least 14 casques of koepckeae, and
they were all similar in size and flat shield-like appearance, which contrasts
with the invariably upright horn of unicornis.
In captive individuals of unicornis
at the Santa Cruz Zoo (Bolivia) the upright horn shape was present from a young
age; although the size of the horn increased with age it didn't change shape or
ever resemble the casque of koepckeae.”
Additional comments
from Jacob Socolar: “I am currently in the field without my
notes to say with certainty how many rectrices I observed with white tips on
the Terborgh-Weske transect. It is indeed possible that the bird only
showed 10 rectrices, although that is not my recollection. In any case, a
recent brown-morph specimen from the east slope of the Sira (details
forthcoming) shows cream (rather than white) tips to the rectrices, and clearly
shows such tips on all twelve rectrices (save the outermost, whose tips are
worn off entirely).
“Furthermore, Weske and Terborgh (1971) note the absence of white
tips on four central rectrices. I am certain that the bird I observed was
not missing four central tail feathers, and so there is definitely variability
in the number of white-tipped rectrices in P.
koepckeae.
“Again, I suspect that this information won't substantially affect
the admirably documented case for a split, but I just don't think that the
white tipping on the tail should go down as an "official" diagnostic
character to distinguish between the forms (absence of white seems to be
diagnostic for koepckeae, but
presence of white is not diagnostic for nominate unicornis).”
Comments from Zimmer: ““YES”.
I think that koepckeae and unicornis could be justifiably split on
the vocal differences alone, but the rather extreme differences in casque
shape, even if proven to vary individually in size, provide clinching
evidence. These differences are not
only consistent with the levels of differentiation between other species pairs
of cracids, but they exceed those differences for many pairs of taxa already
treated as specifically distinct.
Should this proposal pass, I would also recommend adoption of the
English names suggested by the authors for the two resulting species.”
Comments from
Stiles: “YES. Differences in voice and casque shape, probably
both important in mate choice, suggest to me that this split is biologically
meaningful.”
Comments from Pacheco: “YES. A partir do exposto, eu sou favorável ao tratamento
sugerido.”
Comments
from Pérez-Emán: “YES. Vocal and morphological
differences argue in favor of this split. It would be interesting to pursue
Jacob’s question on phylogenetic relationships within the genus Pauxi because koepckeae might not be closer to unicornis
(compared to pauxi).”
Comments from Jaramillo: “YES. In particular, the vocal data is what swayed me the most. The further morphological differences are icing on the cake.”