Proposal (562) to South
American Classification Committee
Recognize newly described Thryophilus sernai
Effect on South
American CL: This proposal
would add a newly described species to the list.
Lara et
al. (2012) discovered a new wren from the dry Cauca River Canyon, an
inter-Andean valley in Antioquia, northwestern Colombia, which is formed by the
northern sectors of
the Western and Central cordilleras and is enclosed at is mouth
by the rainforests of
the lower Cauca basin (Nechí Refuge). We described this population as a
new species. Our proposal is based on an integrative study in the Thryophilus group, including the results
of a comparative study of the distribution, vocalizations, morphology, and
phylogeny. The details can be found in the paper, but in short, we found the
following:
- Plumage
and morphology: The new species is similar to T. rufalbus but is paler overall and more cinnamon-brown rather
than rufous. It is also similar to T.
nicefori, but the latter differs in having a darker, colder brown
upperparts with faint black barring on the dorsum and upper tail-coverts. The
barring of the wings and tail differs decidedly among these three species and
not so much among different subspecies of T.
rufalbus (Fig. 1). Thryophilus sernai
is smaller in body mass and wing length than both T. rufalbus (all subspecies) and
T. nicefori; it has a smaller bill than T.
nicefori, and its tail tends to be longer than T. rufalbus.

Figure 1. Dorsal view
(left to right) of: Thryophilus nicefori,
Thryophilus sernai sp. nov., T.
rufalbus cumanensis and T. rufalbus
minlosi.
-
Vocalizations: The new species fall within the “Thryophilus” vocal group identified by Mann et al
(2009). Recordings of the new species can be examined in the Auk website or in here. Relative
to other taxa in Thryophilus, the new
wren’s songs have a richer repertoire of syllable types, shorter trills, lower
number of trill syllables, a distinctive terminal syllable with more
modulations, and higher spectral frequencies. Although there was some overlap
among Thryophilus taxa as expected in
their “acoustic space” characterized by multivariate analysis, songs of T. sernai are distinct but more similar
to those of nicefori and most
distinct from T. rufalbus. Two
discriminant analyses revealed statistical significant differences in songs
traits of T. rufalbus, T. nicefori, and the new taxon.
Valderrama et al. (2007, 2008)
previously documented the vocal behavior of T.
nicefori and demonstrated its vocal distinctiveness in relation to T. rufalbus.
-
Genetic differences: A phylogenetic analysis based on mitochondrial
DNA (cyt-b) sequence data confirmed
that the new taxon is a member of the genus Thryophilus,
and that it is evolutionary isolated from all other taxa in that genus
(Lara et al. 2012). Sequence
divergence of T. sernai averages the
following values in comparison to other taxa: T. nicefori (3.8%), T. r. castanonotus (3.8%), T. r. minlosi (3.6%), T. r cumanensis (2.7%), T. rufalbus rufalbus (5.4%), and T. rufalbus ssp. (3.7%). The new species, T.
nicefori, and T. rufalbus taxa
(except the last two) are reciprocally monophyletic (Lara et al. 2012, C. D. Cadena et
al., in prep.).
Recommendation:
Lara et al. (2012) concluded that the
population from the dry Cauca Canyon in Antioquia, Colombia, was an unnamed
taxon that merited species rank. The new species has unique vocal, plumage,
morphological traits, it is evolutionarily isolated from closely related forms,
and it occupies a small range, ecologically and geographically separated from
subspecies of T. rufalbus (Fig. 2).
Although reproductive isolation has not been assessed directly, some or all of
the divergent traits of the new species are likely involved in territory
defense, courtship, and mating, and given the current species-level taxonomy of
the group, which recognizes T. nicefori at the
species level, we believe species rank for the new form is warranted.
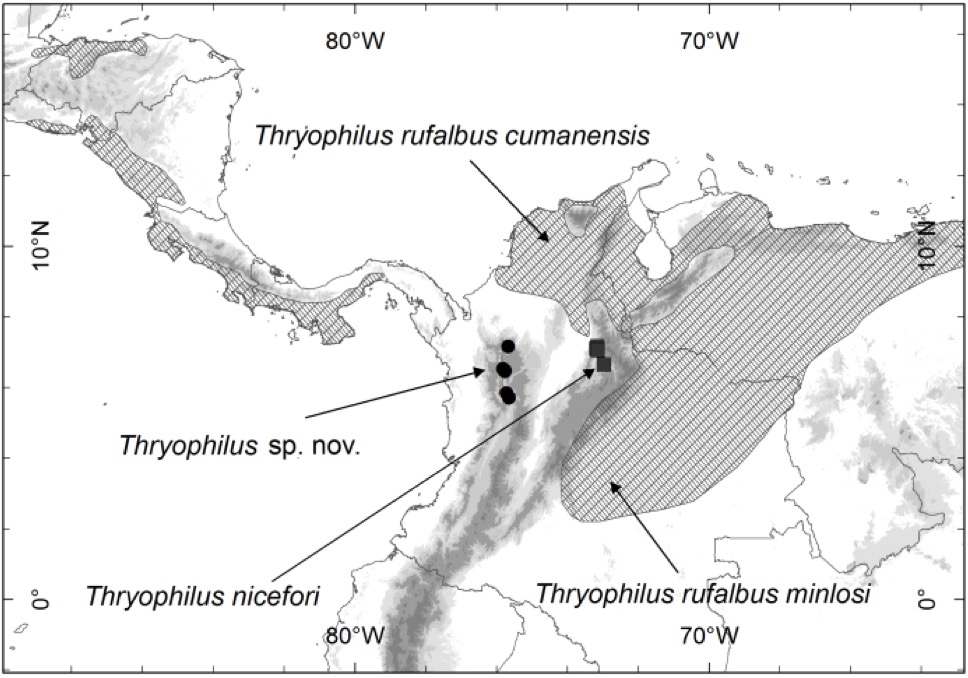
Figure 2.
Distribution of Thryophilus wrens: the new species (black dots), T. nicefori (gray squares), and T. rufalbus (shaded area) in north South America and Central
America.
Moreover, Lara et al.
(2012) stated that: “In comparison to the differences in plumage and song
existing between the two Thryophilus
species pairs known to be sympatric (pleurostictus–sinaloa and pleurostictus–rufalbus), T. sernai is arguably less different
from T. nicefori and T. rufalbus. However, greater
divergence is expected between sympatric pairs of species than between
allopatric pairs (Price 2008). Furthermore, in the context of currently
accepted species limits among members of the T. rufalbus complex (Valderrama et al. 2007, Remsen et al. 2012), T. sernai appears to be just as
divergent (or more so) from T. rufalbus
and T. nicefori as these two good
species are divergent from each other.”
We also realize that the widespread and polytypic wren T. rufalbus as currently defined is not
monophyletic and likely comprises multiple species and is in need of a formal
taxonomic revision. With that analysis pending, however, we believe that
recognition of T. sernai is a step
forward in terms of better describing the species-level diversity of this group
of wrens.
References:
Lara, C.
E., A. M. Cuervo, S. V. Valderrama, D. Calderón-F. and C. D. Cadena. 2012. A
new species of wren (Troglodytidae: Thryophilus)
from the dry Cauca River Canyon, northwestern Colombia. Auk 129: 537–550 – and references therein.
Valderrama,
S. V., J. E. Parra, N. Davila, N., and D. J. Mennill. 2008. Vocal behavior of
the critically endangered Niceforo's Wren (Thryothorus
nicefori). Auk 125:
395–401.
Carlos E. Lara, C. Daniel Cadena, and Andrés M.
Cuervo, October 2012
Comments from Remsen: YES. I
reviewed the paper at pre-publication stage and strongly support the
conclusions. As the authors noted, there
may be additional problems in rufalbus,
but that should not affect the decision herein.”
Comments
from Zimmer:
“YES. The extent of the
morphological and vocal differences would appear to be consistent with
species-level recognition, especially when compared against the “yardstick” of
differences between other taxa currently ranked as species within the genus. Genetic distances between sernai and other congeners are also
consistent with such a ranking, and, recognition of sernai as a distinct species makes sense biogeographically.”
Comments from Stiles: “YES. Morphology,
vocalizations, and genetics are in accord with species status for sernai, regardless of what finally
happens in rufalbus (where minlosi at least might merit species
status).”
Additional comments from T. Donegan: “We recently assessed this species for the Colombian checklist (reference and link below). The text of our conclusions is set out below:
‘Antioquia Wren Thryophilus sernai
Recently described from the Cauca valley in Antioquia by Lara et al. (2012) as a species. This is clearly a new taxon, and we congratulate the discoverers. Any decision to assign it species rank (separately from allopatric Niceforo's Wren T. nicefori and T. rufalbus) at the present time is moot but we follow Lara et al. (2012)'s approach on account of this being a plausible long-term treatment. The new species is illustrated in Fig. 3 and its distribution is shown in Fig. 11. There are specimens (including the type specimens) and published sonograms and photographs of T. sernai from Colombia, so it can clearly be considered "confirmed" in the country to which it is endemic.
‘Recognition of this species has proved to be one of the more controversial issues considered in this series of annual papers on the Colombian checklist. In discussing species limits, Lara et al. (2012) considered that "it is likely that T. sernai has differentiated from T. nicefori and T. rufalbus to the point that they would behave as reproductively isolated units should they come into contact" citing differences in morphology, mtDNA and song. They claim in the diagnosis section that the new species is "distinctive in nine acoustic variables" and that it has a "richer repertoire of syllable types, shorter trills, lower number of trill syllables, a distinctive terminal syllable with more modulations, and higher spectral frequencies". However, there is no data available that would suggest that sernai is diagnosable to the usual 97.5% benchmark (Isler et al. 1999) used for supporting species rank determinations vocally. Their vocal "diagnosis" is based on the Kruskal-Wallis test (in Lara et al.'s table 3), which determines the likelihood that data sets come from populations with different medians, but says nothing about the extent of differences or diagnosability. No standard deviation data is presented, so there is no way of reverse engineering the data for these purposes. In studies of other taxa, pairwise mean differences have sometimes been found consistent with miniscule differentiation and low levels of diagnosability (e.g. Donegan 2012a). There is c.91% differentiation and considerable overlap based on recorded values in multivariate space (their figure 6) suggesting that voice is not diagnostic. The authors' claim of shorter trills is not borne out by the illustrations in the paper (2, 5 or 9 notes in the trills in Fig. 3 for sernai, versus 2-8 in other taxa). The claim of a richer repertoire of syllables is subjective. Differences in modulation of the final note and overall maximum acoustic frequency are true of some but not all examples of songs in their figure 3, so again are not diagnostic. Song can be learned in oscines such as wrens, so the possibility that differences may be cultural and perhaps could be eliminated by learning if populations were to come into contact cannot be easily dismissed. No mention is made of whether sernai responds to playback of related species or how.
‘Lara et al. (2012) note that the rufalbus group requires revision but also consider that: "the paraphyly of species is an expected outcome of speciation processes in which differentiation occurs in peripheral populations". There is at least one documented instance of this phenomenon in Troglodytidae (Troglodytes cobbi: Campagna et al. 2012). However, T. cobbi is strikingly different in its ecology (absence from human modified habitats) to T. aedon, whilst T. sernai is a differently marked version of T. rufalbus / nicefori in a different dry valley. It has a more proximate distribution to rufalbus and nicefori than T. cobbi does to T. aedon. New taxon sernai is less differentiated in its mtDNA than some other named populations in the rufalbus group are from one another (2.5-3.5% between nicefori, sernai and proximate rufalbus; compared to 6.8% between nominate T. rufalbus and subspecies castonotus). In conclusion, it seems implausible that a rational treatment for the rufalbus group involves only T. nicefori and T. sernai being afforded species rank. At least, nominate rufalbus and its relatives would also appear to need splitting from the southern rufalbus taxa.
‘Despite these concerns, we recognise T. sernai on account of its broadly similar levels of vocal
differentiation from rufalbus to that
of nicefori, which is historically widely recognised as a species and shows
similar vocal differentiation from other taxa (Valderrama et al. 2007). Long-term, splitting sernai, nicefori and some other rufalbus
taxa would seems a reasonable approach. In molecular phylogenies, sernai (like T. nicefori) is nestled within T.
rufalbus and is similarly differentiated to T. nicefori. Moreover, it would be a questionable outcome to
see a potentially threatened taxon like this, with a unique distribution go
unprotected whilst an open-ended taxonomic revision takes place. A revision of species limits in the T. rufalbus group as a whole is urgently
called for however.’
“For such an
accessible population, Lara et al. (2012)'s lack of mention of playback
experiments seems anomalous and raises most questions. The results of playback studies were key in
affording species rank to Henicorhina
negreti, the only other recently described wren species occurring in
western Colombia. This new species was
described by Salaman et al. (2003), a paper highly relevant to the sernai description but not cited. It seems unimaginable that the authors would
not have played sernai an MP3 of nicefori and/or rufalbus given the widespread and low-cost use of Ipods and similar
devices for playback nowadays and easy availability of recordings of these
other species. Did sernai respond? T. rufalbus has never been that difficult
a bird, and now T. nicefori is also
accessible in a few known localities. Do
they respond to sernai? How? Is this relevant to species limits?
“There would not
seem any reason of principle as to why a species should not be recognised in a
paraphyletic group like this, even if the description increases overall
paraphyly, where the treatment is defendable as a long-term approach and
differentiation of the study taxon is consistent with that of other
species. The following might be a less
speculative justification: "The
paraphyly of species has been found in some better-studied groups and is an
expected outcome of historical processes for nomenclature and taxonomy in
groups which only recently have been subject to vocal and molecular
investigations".
“Reference: Donegan, T.M., Quevedo, A., Salaman, P. &
McMullan, M. 2012. Revision of the status of bird species occurring or reported
in Colombia 2012. Conservación Colombiana 17: 4-14.
http://www.proaves.org/proaves/images/RCC/Con_Col_17_1-14_Actualizacion_Listado.pdf”
Comments from Cadena: YES (I am
a coauthor of the description). Regarding the "anomaly" noted above,
I do not know if people have played the song of nicefori and rufalbus to sernai; I only saw the bird in life for
the first time last month. I bet people probably have, however, and considering
that nicefori responds to songs of rufalbus and that wrens may often be
interspecifically territorial (good published examples in Thryothorus and Henicorhina),
I bet that sernai likely responds.
What would that mean? This is something one cannot tell without carefully
designed reciprocal experiments (its not simply a matter of taking an iPod once
to the type locality), which someone should probably carry out at some point
when a much-needed revision of the whole rufalbus
complex is undertaken. I will leave it to readers and to other committee members
to evaluate whether the additional criticisms are valid. I stand by our
conclusions that, taken together, and relative to the existing species-level
taxonomy of the group, sernai is best
ranked as a species-level taxon.”
Comments from Nores: “YES. Vocalizations, morphology, and especially the genetic differences match the species status for this taxon.”
Comments from Robbins: “YES. All data support the recognition of T. sernai as a species.”
Comments from Stiles: “YES. Recognizing sernai as a distinct species (at least pending a general in-depth
study of the entire rufalbus complex)
seems the best option.”
Comments
from Pacheco:
“YES. I also agree that treatment as a
species is the best choice for this complex.”