Proposal (624) to South American Classification Committee
Treat Cacicus
flavicrissus as separate species from Cacicus
cela
This
proposal is to split Cacicus flavicrissus
(P. L. Sclater, 1860) together with
Cacicus vitellinus (Lawrence, 1864) (both as the Saffron-rumped Cacique)
from Cacicus cela (as Yellow-rumped
Cacique). Recent authorities have
treated flavicrissus and vitellinus as subspecies of Cacicus cela, notably Howard & Moore
(1994), Dickinson (2003), and Fraga (2011).
They are as follows:
vitellinus from central Panama south to northern and
central Colombia: east to Santa Marta Mountains and south to Tolima on western
Slope of Eastern Andes;
flavicrissus west Ecuador south to
northwestern Peru (Tumbes).
cela
Colombia: eastern lowlands east of the Andes, including north to coast
near Santa Marta Mountains, most of Venezuela, Guyana, Surinam, French Guiana,
south to east Ecuador, eastern Peru south to lowlands of Santa Cruz, and Brazil
(Amazonia from northern borders south to Mato Grosso do Sul).
However,
one should note that Fraga (2011: 749),although he followed the precedent of
three subspecies, stated: “Races form two distinct groups,
separated by the Andes. Western group (vitellinus
and flavicrissus) possibly represents
a separate species., differing from single-species “cela group” in plumage, colour and voice.”
Given
that the two groups are separated for most of their range by the Andes, gene
flow between them must have ceased some millions of years ago. In terms of
plumage, Fraga (2011: 749-750) stated: Male [cela] is mostly black with slight bluish
gloss; lower back, rump, uppertail-coverts and read abdomen yellow; various
amounts of yellow in upper-wing, affecting great coverts and some medium
coverts; uppertail mostly black from above with yellow at the upper edges,
undertail yellow, iris blue, bill ivory; legs blackish.” He continues (2011: 750): “Race
vitellinus has yellow parts of
plumage richer, more orange-yellow than [cela],
also wing patch smaller, bill greyish at base; flavicrissus is very like previous, but smaller.”
In
terms of voice, Fraga (2011:750) mentions some striking differences: ”[Cela] is an outstanding mimic of many
birds, including parrots,…woodpeckers,…,many passerines and others, even of
mammals. Its variable song often starts with a harsh double-note,”k’wek-k’wok”
followed by imitations. Males of race vitellinus
not known to mimic but still have varied song repertoire.” Clear differences between calls is clear from
the following spectrograms, and the calls themselves can be heard on my website
(worldbirdinfo.net):
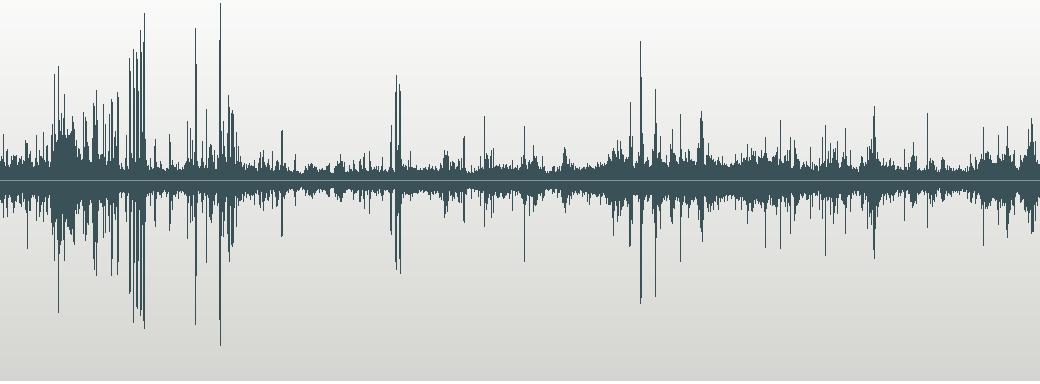
Spectrogram of Call of
Saffron-rumped Cacique Cacicus
flavicrissus vitellinus
Darien, Panama. Thanks
to Linda Macaulay
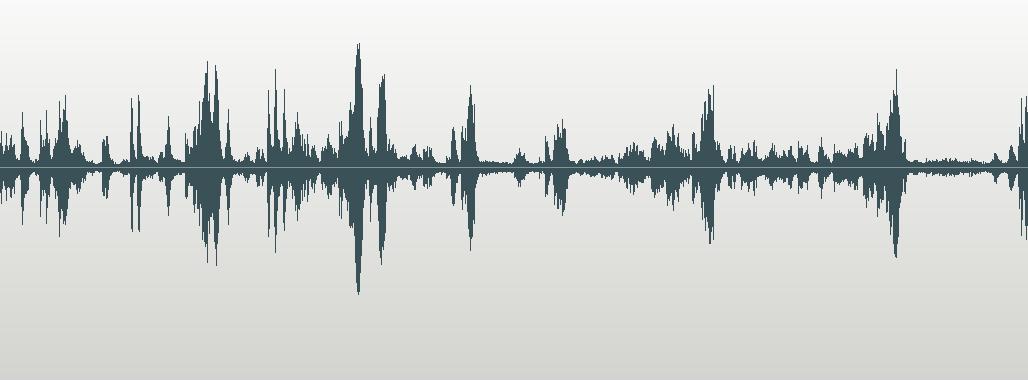
Spectrogram of
Saffron-rumped Cacique Cacicus
flavicrissus flavicrissus. Guayas, west Ecuador

Spectrogram of Call of
Southern Yellow-rumped Cacique Cacicus
cela. Call taped at Cojedes Venezuela by Ted Parker
They
similarity of the calls of the two taxa placed in Saffron-rumped Cacique should
be obvious, as should the difference of both from that of the Yellow-rumped
Cacique.
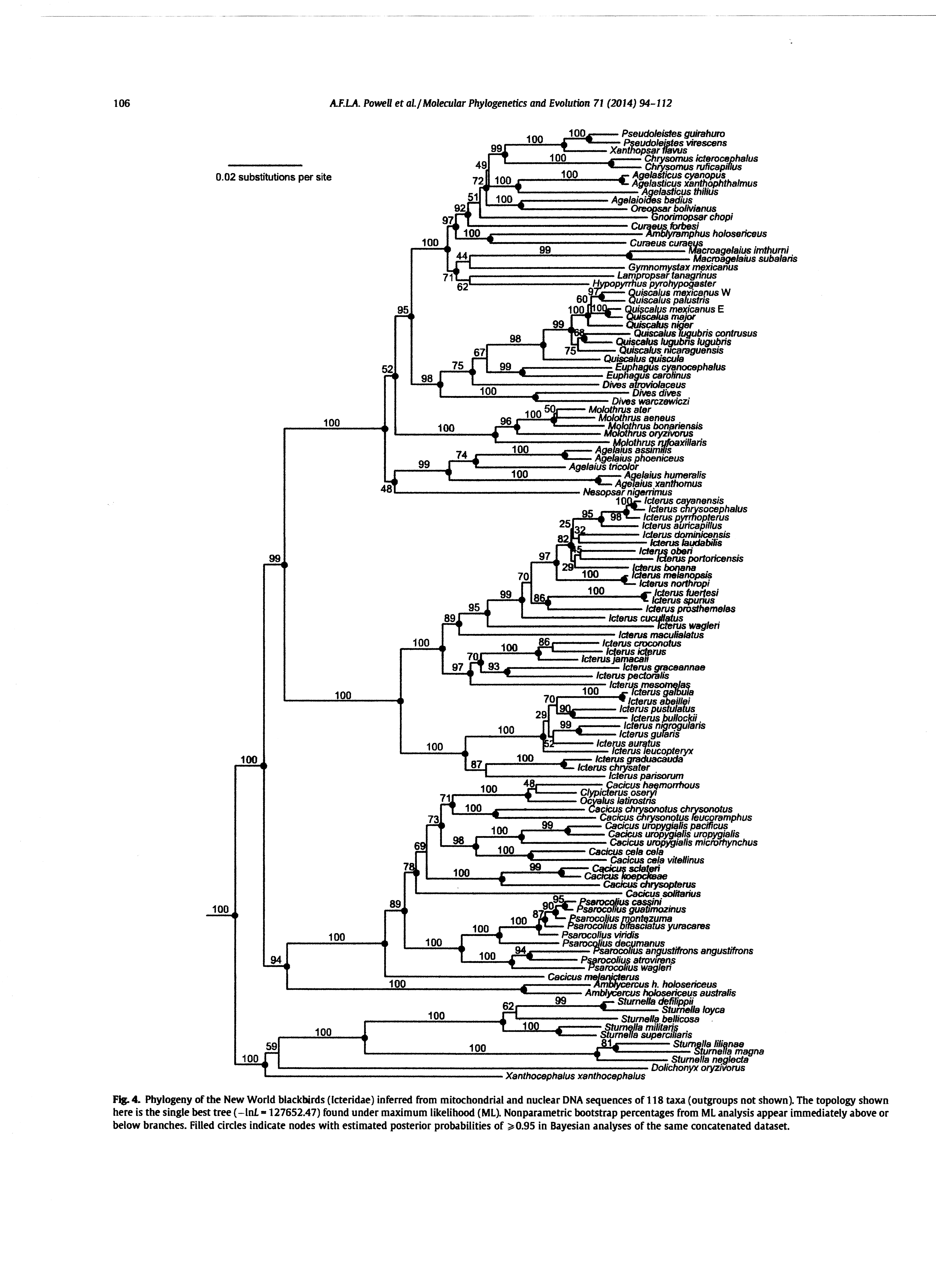
We
may finally turn to genetic evidence. In Figure 4 of Powell et.al, (2014:106)
the node uniting Cacicus cela cela
and Cacicus cela vitellinus is relatively
deep. It is deeper than the node uniting
Cacicus sclateri and Cacicus koepckeae. It is also deeper
than the clade uniting Cacicus haemorrhous
and Cacicus (was Clypicterus) oseryi.
Looking further afield, it is deeper than the nodes uniting many species of Psarocolius.
I
believe that this body of information justifies the split of Cacicus flavicrissus flavicrissus (P.L. Sclater,
1860) (with subspecies Cacicus flavicrissus vitellinus
Lawrence, 1864), Saffron-rumped Cacique, from the now monotypic Cacicus cela (Linnaeus, 158),Yellow-rumped
Cacique.
References
Powell, Alexis F.L.A., Barker, F. Keith, Lanyon, Scott M., Burns,
Kevin J., Klicka, John, Lovette, Irby J.
(2014) A comprehensive species-level molecular phylogeny of the New
World blackbirds (Icteridae) Molecular Phylogenetics
and Evolution 71: 94-112.
Fraga, R. M, (2011)
Family ICTERIDAE (New World Blackbirds) in Del Hoyo, J., Elliott, A., D. Christie,
D. (eds), Handbook of the birds of the world,16, Tanagers to New World
Blackbirds, Barcelona: Lynx Edicions, pp.684-810.
John
Penhallurick, Feb. 2014 (revised March 2014)
__________________________________________________________________________________________________________________________________________
Comments
from Cadena: “NO. For
one, there are no published analyses of vocalizations with proper sample sizes,
sufficient geographic sampling, statistics, etc. to back this up. Also, based
on these oscillograms (note they are not spectrograms) it is hard to visualize
how different songs really are. Also, see my comments on proposal 616 regarding
genetic distance.”
Comments
from Dan Lane: “Although
I am actually of the mind that flavicrissus (presumably also including vitellinus)
should be separated from cela at the species level, I am sorry to say
that the present proposal does not offer the information necessary to support
such a move. First of all, in Powell et al. (2013), C. c. flavicrissus
is not included among the taxa compared in the tree constructed. This, and the
caveats that Van gave in proposal 616 (that a family-wide phylogenetic paper
with only one or two samples per taxon taken from distant points in their
distributions, etc.), means that Powell et al. (2013) is not a work that can be
used to support a taxonomic change of this nature. Secondly, listening to
Xeno-canto, I found a cut of C. c. vitellinus that seems to include
imitations (see http://www.xeno-canto.org/47092,
in which I hear imitations of Campephilus vocalizations). Certainly,
that doesn’t mean that these taxa shouldn’t be split, but it does mean that the
argument using strictly the presence/absence of imitations or geographic voice
variation needs to be more thorough (not to mention, use larger samples and
homologous vocalizations and take into account the fact that these are
oscines!). The three waveforms (not spectrograms) posted in the proposal are
fairly useless in making a case here as the source recordings are not available
on <http://worldbirdinfo.net/Pages/BirdMediaView.aspx?BirdID=40718&Source=%2FPages%2FBirdsSearch.aspx%3FBirdField%3D0%26BirdSearch%3Dcacicus%2520cela>
as far as I can tell, which also severely hampers their usefulness in making
any argument. Finally, according to Hilty and Brown (1986, Birds of Colombia), C.
c. cela is found north of the Colombian/Venezuelan Andes (so the Andes are
not the barrier between the two groups that the proposal makes them out to be)
to the north flanks of the Santa Martas, and at the same time, C. c. vitellinus
is found to the western and southeastern flanks of the same range. With
anthropogenic habitat conversion perhaps allowing for the expansion of these
disturbed-habitat-loving species to spread further, it may be possible to use
this region as a natural laboratory to see how the two interact (if they
actually come into contact) with one another. A more densely sampled molecular
study of these three taxa, with perhaps an associated comparison of a larger
sampling of vocalizations (carefully screened to compare only homologous
vocalizations) may be able to make the argument that was attempted in the
present proposal. As it is, I am afraid the evidence provided here is cannot
support the case.”
Comments
from Remsen: “NO. Although I thank John for provoking
discussion on this an illuminating the need for additional data and analyses,
the data at present are inadequate for changing the classification, for reasons
noted above by Daniel and Dan.”
Comments
from Stiles: “NO.
Again, I think that a good case could be made for this split, but the data
presented here are not sufficient to justify it at present.”
Comments from Nores: “NO, for the reasons given by Lane
and Daniel The cis- and trans-Andes distribution of the two taxa is not
correct. Moreover, it is a complicated procedure to use genetic distance to
assess species status. I don’t consider that genetic data are (yet) useful in
evaluating species limits.”
Comments
from Zimmer: “NO, for
reasons elaborated by others. A more
thorough, quantitative vocal analysis is required.”
Comments
from Jaramillo: “Comments from Jaramillo: “YES. Again, there are problems with this proposal as others
note, but some details regarding song differences are clear and known. Cacicus cela does not mimic during all
song types, but one specific song type known as the harsh song, which is aimed
mainly at rival males. I could not hear the possible mimicry that Dan mentions,
but he has a way better ear than I do! Even if mimicry is done at times by flavicrissus, it does not do it
consistently and as part of a separate song type as cela does. Apart from the mimicry issue, the song structures are
very different between the two species, and behavioral papers have published
information on songs, which I summarized below in our Icteridae book. I am sure
more is known now, but I have not kept up.
“Perhaps most
importantly is that the ranges of these two taxa practically touch, they are
not separated by the Andes as noted in the comments made by a couple of folks.
Yet there are specimens from these areas, and there is no evidence of
intergradation. To me there are several important points here 1) genetics
suggest a deep division between the two 2) songs differ in structure, quality,
and perhaps cela has a song type
entirely missing from flavicrissus. Calls are also consistently different. 3)
Ranges nearly touch, no evidence of intermediates. 4) Other taxa of caciques
and oropendola currently considered separate species are less distinct than
these two.
“Below are
parts from our Icteridae book, which is not a common one on the shelves, but
summarizes a lot of what was known then regarding these birds”
VOICE The repertoire of vocalisations in this species
is huge making a summary of their vocalisations somewhat futile. In addition,
C. c. cela is an adept mimic. Song: The primary song (‘cela song’) is usually
made up of four syllables, the first of which is a screech typical of
Yellow-rumped Caciques. The following notes are variable. It lasts under two seconds
in duration. Each colony has a song (or several songs) specific to the colony
and all males conform to these songs while at the colony. The colony song
changes from year to year, and sometimes within a season. While in full display
another song is used (‘harsh song’), this is more complex and continuous, not
discrete. This longer song that can last up to 20 minutes includes many loud
whistles and it is here that this cacique (form C. c. cela) will incorporate
mimicry of local birds, frogs and assorted noises. The harsh song is extremely
variable, but includes the ‘tchak’ notes typical of this cacique. The loud
volume and varied nature of the song may be its most distinctive features.
Females rarely sing the ‘cela song’ but never sing the ‘harsh song’. It appears
that ‘harsh song’ is aimed mainly at other males and has some aggressive or
threatening significance, while ‘cela song’ is correlated with slight alarm and
may function to advertise the presence of the male to females (Feekes 1981).
The form vitellinus
is not known to mimic or to produce mechanical sounding noises. The
vocalisations of vitellinus are said to be conversational and musical series of
notes: ‘wick-a-weo’ ‘char-che-ar’, ‘chut-chu-chu’. Also gives a more muffled
song, ‘Aaaghh-a-whaaghee’, or ‘cluuk-cluuk-whaaagooo’. Males of vitellinus sing
five to eight songs types; all males in a colony share the same song types. The
number of shared songs between colonies is directly proportional to the
distance between colonies, nearby colonies share more song types. Dialects are
stable during a breeding season, but some songs change gradually over the
months. Less than half of the song types are present in a recognisable form
during the next breeding season. Some new songs (17%) are introduced by dispersing
males from other colonies, but for the most part new songs are derived from old
songs (45%), the rest are of unknown origin (Trainer 1989). An analysis of song
types of vitellinus in Panama discovered that of the seven types specific songs
were associated with certain behaviours. One song was more likely to be sung by
males in groups, while another was sung by lone males, another by flying males,
another when supplanting another male while the rest were not so closely
associated with specific contexts (Trainer 1987). There appear to be consistent
rules that govern the sequence of song types given by males at the colony.
Social interactions as well as social context appear to influence (or
determine?) the organisation of song types at the colony (Trainer 1988). The
form flavicrissus of the Pacific Coast is also not known to be a mimic. Calls:
The common call of cela is a repeated harsh ‘tchak’ or ‘chaaak’, uttered more
loudly and frequently when alarmed. Females utter a rough ‘rrrrrrr’, particularly
when behaving aggressively. The subspecies vitellinus gives a standard icterid
‘chuk’.
GEOGRAPHIC VARIATION Three subspecies are recognised
which fall into two groups: vitellinus (Saffron-rumped, or Lawrence’s Cacique)
and cela (Yellow-rumped Cacique). [NOTE of correction here – I am not
sure why I used the name vitellinus,
if separated I gather that flavicrissus
has priority] It is clear that there are
consistent morphological, behavioural and vocal differences between these two
groups and it may be determined that they are best separated as two different
species in the future. The two groups almost come into sympatry in N Colombia,
but no intermediate specimens are known.
Yellow-rumped
Cacique C. c. cela is found east of Andes. This is the most widespread
subspecies, found throughout the tropical lowlands of South America east of the
Andes as far south as C Bolivia and E Brazil. In Colombia, cela is found along
the coast of Santa Marta, and comes quite close to the range of vitellinus.
This population shows no evidence of intergradation towards vitellinus (Todd
and Carriker 1922). Cela is described above.
The
Saffron-rumped Cacique is found west of Andes and is comprised of two
subspecies. The race C. c. vitellinus is found from the Canal Zone in Panama
east into W Colombia (south to the Rio Salaqui) to the western and southern
base of the Santa Marta mountains, south to the middle of the Magdalena Valley.
It differs from cela in being larger, with a bill showing a more strongly
curved and broadly based culmen. The yellow of the body is a deeper colour,
more orange, than on cela. As well the yellow on the coverts is much less
extensive, being restricted to the innermost greater coverts. The yellow on the
base of the tail is much less extensive, only covering the basal third or less
on the outer rectrices. Wing Formula: P9 < P8 < P7 > P6; P9 » P5; P8 - P5 emarginate.
The other race in this subspecies group, C. c. flavicrissus occurs in
the Pacific lowlands of Ecuador from Esmeraldas south to NW Peru (Tumbes).
There is a gap in range between flavicrissus and vitellinus along the Pacific
Coast of S Colombia. This form is like vitellinus in having less yellow on the
tail than cela, but it has a larger yellow wing patch, like cela. It is smaller
than vitellinus, has less of an orange colour on the rump, and a smaller bill
which has a plumbeous coloured base. Wing Formula: P9 < P8 < P7 > P6;
P4 < P9 < P5; P8 - P5 emarginate.
Comments from Robbins:
“NO. Dan does an excellent job of summarizing all the shortcomings of our
knowledge of the cela complex. Until additional data are forthcoming, I see
no unequivocal evidence for treating these taxa as separate species.”
Comments from Pacheco: “NO. Compartilhando da opinião preponderante, aguardo uma
publicação que faça a análise das vocalizações, a partir de amostras em número
apropriado e bem distribuídas geograficamente para uma decisão consistente da
situação.”