Proposal (628) to South
American Classification Committee
Reassign
species currently placed in Myrmeciza
into 12 genera
Effect
on SACC:
Species currently placed in Myrmeciza
will be assigned to Myrmeciza and
eleven other genera, five of which (Akletos
Dunajewski, Myrmelastes Sclater, Myrmoderus Ridgway, Myrmophylax Todd, and Sipia
Hellmayr) are resurrected, and six of which (Ammonastes, Ampelornis, Aprositornis, Hafferia, Poliocrania,
and Sciaphylax) are newly described.
Species currently placed in Schistocichla
will be reassigned to Myrmelastes.
Background: The history of Myrmeciza is characterized by decades of uncertainty and controversy
as outlined in Isler et al (2013). Recent authors have consistently expressed
doubt that Myrmeciza represented a
monophyletic group, but until recently the basis for a revision was
unavailable.
New Information:
Results from a subset of taxa from a densely sampled molecular phylogeny (218
of 224 species) of the Thamnophilidae (Isler et al. 2013, Figure 1) provided
high support that Myrmeciza is not
monophyletic and that its members are located in three of the five tribes of
the subfamily Thamnophilinae proposed by Moyle et al. (2009). Morphological,
behavioral, and ecological character states of Myrmeciza species and their closest relatives within each tribe
were compared to establish generic limits. Finally, results were arrayed to
test for consistency and provide a benchmark for replication. Additional
perspective was provided by examining recommendations in a time-calibrated
species-level phylogeny of the entire family (Bravo 2012).
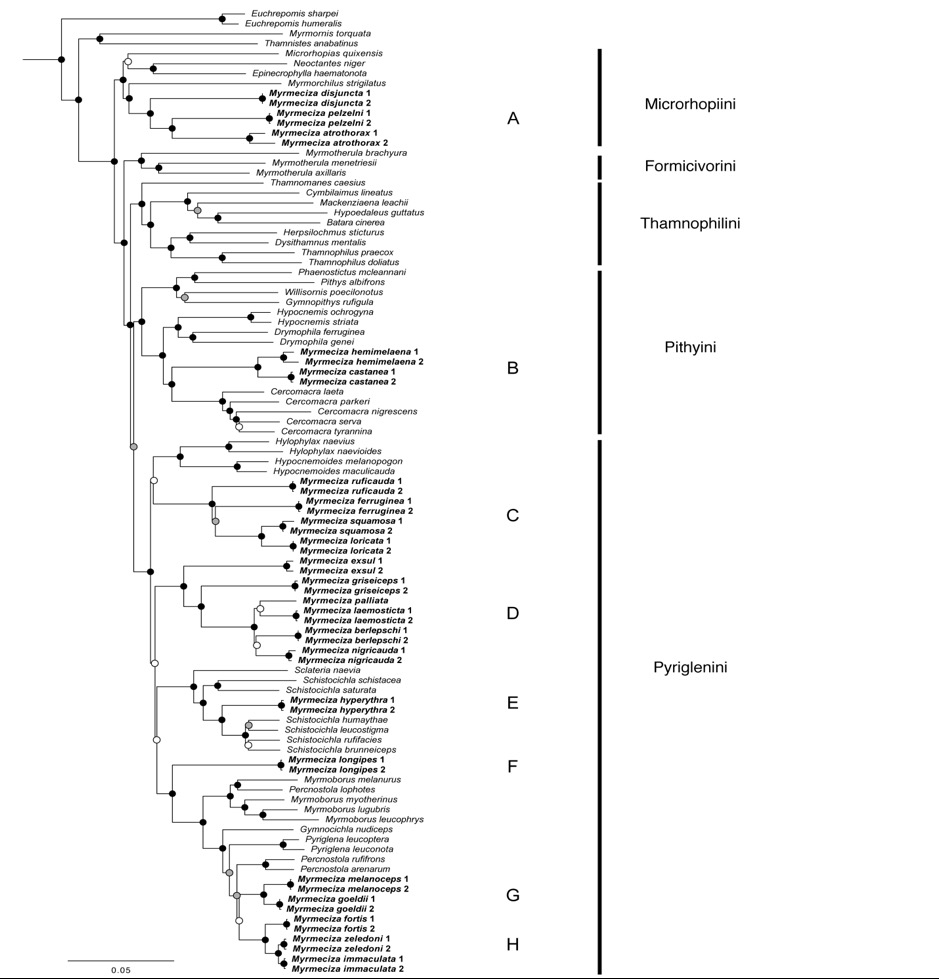
Figure
1. Maximum-likelihood tree of a subset of the Thamnophilinae (Myrmeciza species are emboldened). The
color of the circles at nodes indicates bootstrap support values, > 70%
(black), 50-70% (gray), < 50% (white). This figure is Figure 1 from Isler et
al. (2013):
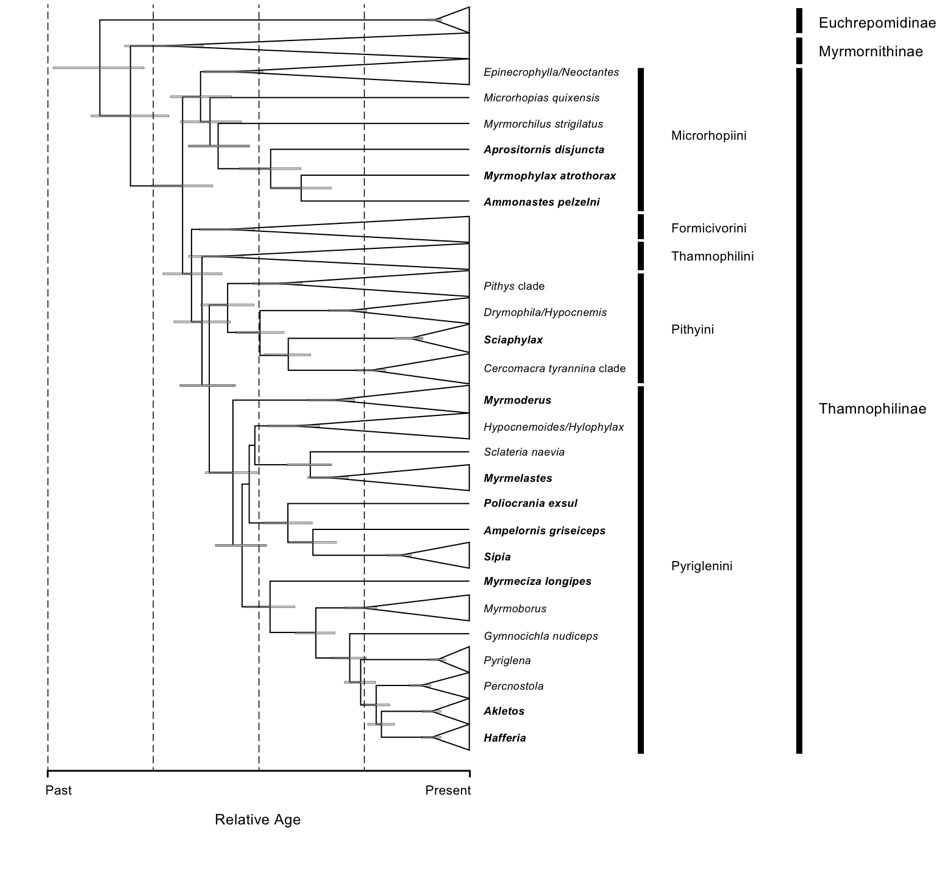
Figure
2. Time-calibrated tree showing relative ages of former members of Myrmeciza. Bars at nodes indicate the
95% highest posterior density for the inferred divergence time estimates. This
is Figure 3 from Isler et al. (2013) with the Akletos revision.
Proposals:
Because
the number and type of supporting traits is greater for some generic
recommendations than others, we have subdivided this proposal into seven parts
and have provided alternatives that the committee might wish to consider for
some of them. More extensive discussion of the basis for our recommendations
can be found in Isler et al. (2013). Table 1 below provides a summary of
recommended genera and species and suggests interim placement of each genus in
the current SACC sequence pending a comprehensive revision of the Thamnophilidae.
Proposal
628A.
Recognize the genera Myrmophylax, Aprositornis, and Ammonastes. Although
we were anxious to avoid monotypic genera, study results required recommending
a number of them. The three "Myrmeciza" species located in the
tribe Microrhopiini ("A" on Figure 1; Figure 2) differed
diagnostically from one another and from their closest relative, Myrmorchilus strigilatus, by multiple
traits, more numerous than well established thamnophilid genera (see Isler et
al. 2013, Table 4). Any other treatment would result in non-diagnosable
heterogeneous genera. Relative crown and stem ages of such genera would be
comparable to those of complete tribes of the Thamnophilinae (Figure 2)
Proposal
628B.
Recognize the genus Sciaphylax. The
same is true for the comparison ("B" on Figure 1; Figure 2) between
the pair of "Myrmeciza"
species located in the tribe Pithyini with their closest relative, the "tyrannina clade" of Cercomacra which has recently been
recognized as a distinct genus, Cercomacroides
(Tello et al. 2014).
Proposal
628C.
Recognize the genus Myrmoderus. The
remaining "Myrmeciza"
species are located in the tribe Pyriglenini. Multiple traits distinguish the
four species in this genus, which constituted a well-supported clade in the phylogeny ("C"
on Figure 1; Figure 2).
Proposal
628D (option 1).
Recognize the genera Poliocrania, Ampelornis, and Sipia. The phylogeny identified a well-supported clade
("D" on Figure 1; Figure 2) with exsul
at the base and including griseiceps
and an internally unresolved subclade that includes palliata, laemosticta, nigricauda, and berlepschi. The difficulty is that exsul is similar in morphology and behavior to the subclade, as
suggested earlier by Robbins and Ridgely (1991), but differences in multiple
traits between griseiceps and other
members of the clade indicate that griseiceps
should not be placed in the same genus as others in the clade. If griseiceps is placed in a monotypic
genus, exsul also has to be placed in
a monotypic genus if a paraphyletic genus is to be avoided. Nest architecture
differences between exsul and the
subclade support their separation.
Proposal
628D (option 2).
Recognize the genera Ampelornis for Myrmeciza griseiceps and Sipia for M. exsul, M. berlepschi, M. nigricauda, M. laemosticta, and M.
palliata. Not recommended. This option creates a paraphyletic genus.
Proposal
628D (option 3).
Recognize the genus Sipia for Myrmeciza exsul, M. griseiceps, M. berlepschi,
M. nigricauda, M. laemosticta, and M.
palliata. Not recommended. This option creates a genus diagnosable only
geographically that includes species whose traits are inconsistent as compared
to other thamnophilid genera.
Proposal
628E (option 1).
Recognize the genera Sclateria and Myrmelastes. The phylogeny identified a
well-supported clade ("E" on Figure 1; Figure 2) with Sclateria naevia at the base and
including Myrmeciza hyperythra
imbedded within subclades that include Schistocichla
schistacea, S. saturata, S. humaythae, S. leucostigma, S. rufifacies,
and S. brunneiceps. Although not
previously recognized (perhaps because of its larger size), multiple plumage
and behavioral traits of M. hyperythra
are consistent with those of Schistocichla
species. The unique morphological and behavioral characteristics of Sclateria naevia that led to its earlier
placement in a monotypic genus are sufficiently distinct to maintain Sclateria as a monotypic genus.
Proposal
628E (option 2).
Recognize the genus Myrmelastes and
make Sclateria a junior synonym. Not
recommended
Proposal
628F. Maintain Myrmeciza as a
monotypic genus.
Myrmeciza longipes holds a
well-supported position ("F" on Figure 1; Figure 2) at the base of a
clade containing the remaining genera within the tribe Pyriglenini, from which
it differs in multiple traits. Future studies may find that more than one
species is involved.
Proposal
628G (option 1).
Maintain Percnostola and recognize
the genera Akletos and Hafferia. Traits distinguishing genera
other than Myrmeciza in the final
clade are fewer than the preceding clades and almost entirely confined to
morphology, yet the clade includes well-established genera such as Myrmoborus and Pyriglena. The principal challenge involves the Percnostola, immaculata, and melanoceps
subclades ("G" and "H" in Figure 1). A sister relationship
between any of them is not strongly supported so we currently have a polytomy.
Future analysis with additional data can either support: (a) a sister
relationship between Hafferia and Akletos or (b) a sister relationship
between Percnostola and either Hafferia or Akletos. Given the uncertainties, we wanted to avoid lumping Akletos
and Hafferia into one genus that could become paraphyletic. Our
preferred options are either to treat them as separate genera (628G option 1)
or to merge them into Percnostola (628G option 3). With either of these, no
matter what the final topology will be, no taxon can become non-monophyletic.
On the other hand, if Percnostola and
Hafferia + Akletos are placed into two genera (628 option 2), scenario (b)
would render a paraphyletic genus. We recognize that, despite this possibility,
option 2 might be considered a more conservative approach. Given the choice of
placing them in the same genus as the morphologically dissimilar Percnostola or recommending three
genera, the latter was deemed consistent with the morphological distinctions
currently distinguishing other genera in the Pyriglenini. Note that subsequent
to publication, it was brought to our attention that the proposed name for one
of the recommended genera, Inundicola,
was a junior synonym of Akletos, and
a correction has been made (Isler et al. 2014).
Proposal
628G (option 2).
Maintain Percnostola and place Myrmeciza melanoceps, M. goeldii, M. fortis, M. zeledoni, and
M. immaculata in Akletos. Not recommended for reasons discussed under option 1.
Proposal
628G (option 3).
Recognize the genus Percnostola for P. rufifrons, P. arenarum, Myrmeciza
melanoceps, M. goeldii, M. fortis, M. zeledoni, and M. immaculata.
Not recommended. This option creates a genus that includes species whose traits
are inconsistent as compared to other thamnophilid genera.
Proposal
628G (option 4)*:
Recognize Gymnocichla but lump all of
Percnostola, Akletos and Hafferia into
Pyriglena. Not recommended. Same
basis as option 3.
Proposal
628G (option 5)*:
Lump all of Gymnocichla, Percnostola, Akletos and Hafferia into
Pyriglena. . Not recommended. Same
basis as option 3.
References:
Bravo, G.A. 2012.
Phenotypic and niche evolution in the antbirds (Aves, Thamnophilidae). Ph.D.
dissertation. Louisiana State University. Baton Rouge.
Donegan, T. M. 2012.
Geographical variation in Immaculate Antbird Myrmeciza immaculata, with
a new subspecies from the Central Andes of Colombia. Bulletin of the British
Ornithologists’ Club 132: 3–40.
Isler, M. L., G. A.
Bravo, and R. T. Brumfield. 2013. Taxonomic revision of Myrmeciza (Aves:
Passeriformes: Thamnophilidae) into 12 genera based on phylogenetic,
morphological, behavioral, and ecological data. Zootaxa 3717 (4): 469–497.
Isler, M. L., G. A.
Bravo, and R. T. Brumfield. 2014. Inundicola
Bravo, Isler, and Brumfield 2013 is a junior synonym of Akletos Dunajewski 1948 (Aves: Passeriformes: Thamnophilidae).
Zootaxa 3779 (3): 399–400.
Moyle, R.G., Chesser, R.T., Brumfield, R.T., Tello J.G., Marchese,
D.J. & Cracraft, J. (2009)
Phylogeny and phylogenetic classification of the antbirds, ovenbirds,
woodcreepers, and allies (Aves: Passeriformes; infraorder Furnariides). Cladistics, 25, 1–20.
Robbins, M. B., and R.
S. Ridgely. 1991. Sipia rosenbergi
(Formicariidae) is a synonym of Myrmeciza
[laemosticta] nigricauda, with comments on the validity of the genus Sipia.
Bulletin of the British Ornithologists’ Club 111:11-18.
Tello. J. G., M.
Raposo, J. M. Bates, G. A. Bravo, C. D. Cadena, and M. Maldonado-Coelho. 2014.
Reassessment of the systematics of the widespread Neotropical genus Cercomacra (Aves: Thamnophilidae).
Zoological Journal of the Linnean Society 170:546–565.
Morton Isler,
Gustavo Bravo, and Robb Brumfield, April 2014
__________________________________________________________________________________________________________
Comments from Thomas
Donegan: With apologies for another long posting here, I would like to comment on part G of this proposal. The published diagnoses for some of the
proposed genera in this proposal are in several cases not supported by other
published data. Some of these proposed
genera might be better recognised as sub-genera or superspecies. In a separate study of M. immaculata and related species (Donegan 2012) (Isler et al.
(2013)'s "Akletos" and
"Hafferia"), I suggested
that it would be better to lump these:
"… Notably, ellisiana
[a synonym of immaculata]
was
originally described in Pyriglena
by
Sclater (1855) so such a treatment would not be novel. Moreover, M. goeldii shares the bright red eye of Pyriglena and Percnostola rufifrons (others in the clade possess red-brown or rufous to
brownish irides) and all of the Pyriglena
/
Gymnocichla / Myrmeciza clade have fairly uniform black male and brownish female plumages, with
some species having white interscapular patches and wing-covert fringes. Songs
of Pyriglena and Gymnocichla are similarly structured to those of Myrmeciza, involving repeated notes at more or less
the same frequency and with a similar note shape to zeledoni and macrorhyncha (e.g., Fig. 5). "
My recent separate revision of Myrmeciza
contains a lot of relevant data for this group that was not used or cited by
Isler et al. (2013). Isler et al. (2013)
instead rely heavily on various morphometric data based on a smaller sample,
but present only averages (and no standard deviations, maxima or minima), so it
was not possible to assess their diagnosis.
I therefore took the dataset on which Donegan (2012) was based, plus my
unpublished mist-net data for Gymnocichla
and Pyriglena, to consider the extent
to which their assertions were supported by my data.
I then considered each claimed diagnostic feature of genera set out in
Isler et al. (2013) to identify those that are unique to their proposed genera
in this clade. "Dome shaped
nests" are mentioned for each genus in diagnosis but do not diagnose
within this group so are not discussed further.
|
Diagnosis |
Comments |
|
Gymnocichla |
|
|
Extensive bare crown
of male. |
Note "Hafferia" have bare skin which is less extensive. |
|
Mostly unmarked black
male |
Shared with male
"Hafferia" (especially immaculata), so not diagnostic. (Males of this monotypic genus have more
extensive white wing covert fringes than immaculata
and its allies.) |
|
Mostly unmarked brown
female |
Shared with female
"Hafferia" immaculata, so not diagnostic. |
|
Differs from Pyriglena in tail and bill lengths |
Supported by my
data. (Not claimed to be diagnostic
against other members of group.) |
|
Pyriglena |
|
|
"brilliant red
eyes". |
Reflects the
vernacular name "fire-eye" but not diagnosable. As noted in Donegan (2012) and the field
guide literature, Percnostola and
"Akletos" goeldii have red eyes. Gymnocichla
have red irides and others in the group have variously dark red, chestnut and
brownish irides. There may be minor
differences in shade of red between the proposed genera. |
|
Large white
interscapular patches. |
Gymnocichla has (smaller) white interscapular patches. |
|
Females with blackish
tails. |
Diagnosable, but note
immature males in a generally female plumage state of immaculata and zeledoni
of "Hafferia" can show
female-like plumages with black tails. |
|
Short bills. |
My mist-net data for
one species in this genus is consistent.
Claimed bill length/total length ratio differences may follow, but
because body length data is so variable depending on specimen preparation
techniques, I do not collate or have such data to compare. |
|
Long tails. |
These overlap in my
data set with the "Hafferia"
(immaculata) group, as noted by
Isler et al. (2013). |
|
Tail length/wing
length ratios, distinguishable from some genera not including "Akletos". |
For tail length/wing
length, my data of "Akletos"
melanoceps is consistent in not overlapping with Pyriglena. However, this
feature is not diagnostic as against all taxa (as noted by Isler et
al.). For M. immaculata macrorhyncha ("Hafferia") this ratio overlaps with Pyriglena. |
|
Percnostola |
|
|
Grey males. |
"Hafferia" fortis is also grey, but darker. |
|
Bill length |
This and other
measurements only asserted as diagnosable against Pyriglena. |
|
Sandy soil habitat
preference. |
Specialisation may be
diagnostic, but "Hafferia"
fortis and some other species in
this clade have a broad ecological range. |
|
"Hafferia" |
|
|
Male black (as several other genera)
or greyish black. |
"Greyish
black" only marginally diagnostic from "grey" for Percnostola. |
|
From Pyriglena (only) in having bare periorbital patch and no
interscapular patch. |
Not diagnosable from
all genera, as noted. |
|
From "Akletos" (only) in "long tail" and "shorter
wings" and "tail length/wing length ratio". |
This difference is
only supported using my data set if particular populations or species are
compared but contradicted by my data when all species are considered. Tail length is diagnostic between "Akletos" melanoceps and
(most but not all) members of the immaculata/zeledoni
group (from among "Hafferia"). However, tail length overlaps substantially
between both species of "Akletos"
and "Hafferia " fortis, so is not diagnostic (Donegan
2012, Appendices 2 &4). Wing length also overlapped for almost all
species in the "Akletos"
and "Hafferia" groups, so
is not diagnostic. Wing/tail ratio
overlapped to a large extent between both "Akletos" and "Hafferia"
fortis so is not diagnostic. These differences presumably result from
Isler et al. (2013)'s smaller sample (245 specimens from 62 species across
all of the broad ex-Myrmeciza and
relatives) compared to mine (283 specimens of just 5 species all in "Hafferia" and "Akletos"). |
|
"Akletos" |
|
|
Diagnosable from other groups
"except Pyriglena and Hafferia" by plumage. |
Non-diagnosable
within clade |
|
Male black |
As M. immaculata, Pyriglena and Gymnocichla. |
|
Female brown or brown and black |
As M. immaculata, Pyriglena and Gymnocichla. |
|
Both male and female lacking wing
covert spots |
As Gymnocichla and "Hafferia" immaculata. |
|
Differs from Pyriglena (only) in having bare periorbital patch (difference
from latter in interscapular markings not mentioned). |
Incorrect: "Akletos" goeldii lacks any visible bare skin. |
|
From Pyriglena in tail and bill length proportions, |
Consistent with my
data but not differing from all other genera in the group in this feature. |
|
From "Hafferia" in shorter tail, longer wing and wing/tail
ratio. |
My data contradicts
this, as set out above. |
“Features unique to a genus
and diagnosable within the Percnostola,
Pyriglena, Gymnocichla, "Akletos"
and "Hafferia" clade are
then only:
Gymnocichla: more extensive bare skin on
head.
Pyriglena: shorter bill, more extensive interscapular
patches.
Percnostola: lighter grey males,
specialism to sandy soil habitats.
"Hafferia": N/A.
"Akletos":
N/A.
“In relation to the latter two genera combined, an "absence of
strong white markings of particular shapes or patterns on the wings" could
perhaps be studied further. Although
claimed biometric differences are not supported by my larger data set, vocal
data in Donegan (2012) show the latter two groups to be diagnosable by the
maximum acoustic frequency and note shape of male songs, multi-note calls and
single note calls.
“Although diagnostic characters are elusive, the general size, songs,
calls, feeding habits, general plumage patterns, nesting and biometrics of all
these proposed split genera are remarkably consistent (see opening quoted
passage). In Donegan (2012, Fig. 5) the
similarly structured songs of Pyriglena,
"Hafferia " and "Akletos" are illustrated. All these proposed genera give songs
consisting of repeated notes of a more or less similar and non-variable shape,
as well as grating "multi-note calls" (Fig. 7 of Donegan 2012) which
vary only in note shape and frequency between species. I would therefore disagree that Proposal 628G
(Option 3) "creates a genus that includes species whose traits are
inconsistent as compared to other thamnophilid genera".
“Based on Isler et al. (2013)'s molecular data, Pyriglena, Percnostola,
"Akletos" and "Hafferia " are also the youngest of
all the clades proposed for species rank.
Gymnocichla is borderline
compared to other genera.
“Defining genera is often a subjective exercise. However, I would personally want to see more than
the above in terms of "real world" differences (or a pressing need to
avoid lumping nice, cohesive genera: cf. the recent Thryothorus proposals and Henicorhina/Cinnycerthia)
to support recognising all these genera in the absence of deeper molecular divergence. John Boyd's "taxonomy in flux"
website independently also lumped "Hafferia
" and "Akletos" for
similar reasons.
“In my view, the most important criteria for genera should be (in order
of importance): (i) assured long-term monophyly and stability; (ii) lack of
damage to existing long-standing treatments; (iii) "real world"
differentiation (morphological, ecological, vocal); and (iv) branch length.
Isler et al. (2013) in this clarified proposal have made good arguments for
Option 2 not being a sensible approach due to risks of paraphyly. Option 1 to my mind involves unnecessary and
unsupported over-splitting: it is based only on lighter grey males and habitat
preferences of Percnostola, claimed
biometric differences that are not supported with a larger data set and
previously unmentioned vocal differences.
At the other end of the spectrum, Options 4-5 would result in
instability through lumping long-recognised genera Percnostola and/or Gymnocichla. As a result, in my view, Proposal 628G (Option 3) (lumping Percnostola, "Akletos" and "Hafferia")
would be the most supportable approach.
We need to change the names of the "Akletos" and "Hafferia"
groups anyhow. This treatment results in
three monophyletic, morphologically diagnosable genera, with branch lengths
more similar to other ex-Myrmeciza. The resulting genera are then all diagnosable
in the real world (Pyriglena:
short-billed, broader interscapular patches; Gymnocichla: uniquely
extensive bare skin; Percnostola:
none of the above).
“I would welcome and support Isler et al. (2013) study and support the
other proposals and contributions in their paper. We are adopting all their other changes in
the soon-to-be published expanded and improved 2nd edition of the
Colombia field guide.”
Comments from Jaramillo: “
A – Yes; B – Yes; C – Yes; D – Option 1
Yes; E – Option 1 Yes; F – Yes; G – Option 1 Yes – this allows for adjustment
if other data come forth.
Comments from
Robbins:
“628a.
Yes, although an argument could be made for treating this clade as a single
genus and if one ascribed to that philosophy and applied consistent treatment
then some of the other proposals would not result in a multitude of
genera. All subjective.
628b.
Yes, but as in above, this clade could be treated as a single genus.
628c.
Yes.
628d.
Yes to option 1.
628e.
Yes to option 1.
628f.
Yes.
628g.
Yes, to option 5. As Thomas Donegan points out, morphological differences are
not that great and if one considers the divergence level and relative time
frame one can certainly argue that this entire clade could be treated as a
single genus. At most, perhaps recognize
Gymnocichla because of the long
branch, but I support treating all these as Pyriglena. Again, all very subjective.”
Comments from Nores: “Note: I am responding to this proposal, based almost
exclusively on the molecular analysis and, except some strictly necessary
cases, avoiding creating new genera.
A. NO. I prefer
to include the three species of this clade in a single genus: Myrmophylax. B. YES.
C. YES D. YES, option 3. E. YES, option 2 F. YES G. YES, option 3.
Comments
from Stotz: “A NO Obviously
these 3 species have to be moved out of Myrmeciza. However, I am okay with recognizing the clade
as a unit. They would be called Myrmophylax,
I think.
B YES
C YES
D Option 3 We lived for a
long time with griseiceps and exsul in Myrmeciza, I think that their differences from the other four can
be survived. While the proposal says that the only thing that diagnoses
the clade is geography, it does seem like geography means something here.
E Option 1
F YES
G Option 3. This is complicated, but
Option 1 creates weakly defined new genera that are basically superspecies. I
am hard-pressed to get rid of Gymnocichla
and Pyriglena. Percnostola,
already seems variable to me, so adding in Hafferia
and Akletos doesn’t offend me.”
Comments
from Pacheco: “A – Yes;
B – Yes; C – Yes; D – Yes to Option 1; E – Yes to Option 1;
F – Yes.
Vocally, it makes sense;
G – Yes to
Option 5 – According to the reasons given by Donegan. Akletos and Hafferia
not seem to be diagnosable a level comparable to other genera.
Comments
from Pérez-Emán: “A
complex proposal that relies somehow in the subjectivity behind how we define a
genus.
628A: YES based on genus diagnosability.
Lumping everything would result in a highly heterogeneous taxon.
628B: YES.
628C: YES. Consistent with morphological
and behavioral characters indicated by Isler et al (2013).
628D: Option 3. I would consider all
species in one genus, Sipia. It makes
sense geographically and we do not have to split this group of related species
into three genera. M. griseiceps is
the most divergent species both morphologically and vocally but these
differences could be considered species level differences. This is a Tumbesian
endemic and many of these restricted species are relatively divergent to their
closest relatives (e.g., Hemispingus
piurae, Basileuterus trifasciatus).
628E: Option 1.
628F: YES.
628G: Option 3. Thomas’ comments on the
morphological variation in this group support this option. However, nodal
support seems to be weak in the phylogenetic tree included in the proposal. If
this is so, Option 5 could be a better option. The original article published
by Isler et al (2013) shows, however, a strong support for the node defining
the clade Percnostola, Hafferia and Akletos.”
Comments
from Areta: “I do not
feel very comfortable with creating many monotypic genera, but based on genetic
divergences shown in the tree, I do not see any other solution. The yardstick
may change, however, if the full phylogeny was shown. I also tend to oppose
overlumping in huge uninformative genera.
I would recommend the following:
A-YES.
B-YES.
C-YES.
D-YES to option
1.
E-YES to option
1.
F-YES.
G-YES to option
3, although I am not uncomfortable with option 5 (I was always struck by the Pyriglena-like vocalizations of Myrmeciza immaculata).”
Comments from Stiles:
“A. YES; certainly disjuncta warrants its own genus, and although I am less
enthusiastic about splitting atrothorax and
pelzelni; branch lengths do support
this as well.
B.
YES, no problem.
C.
YES.
D.
YES to option 1, on the basis of avoiding paraphyly and favoring
diagnosability.
E.
YES to option 1: Sclateria seems
sufficiently divergent to warrant its own genus.
F.
YES. Longipes is definitely an
outlier in this group.
G.
The most difficult proposition. As an
aside, I note that red eyes are not necessarily a diagnostic feature of Percnostola, because the minor subspecies group of rufifrons has gray eyes. I am most comfortable with option 3 (YES),
with option 5 as a second choice.”
Comments from Remsen: “As noted by
others, resurrection and creation of monotypic genera seems the only solution
for making our classification follow the new phylogenetic results. For many decades, the polyphyly of Meyer de
Schauensee’s Myrmeciza has been
predicted by field people, but I don’t think anyone predicted it would be THIS
bad.
“A.
YES for reasons stated in the proposal: lack of morphological diagnosability in
an expanded Myrmophylax and
comparative lineage age as indicated by branch lengths. Those who vote no on A need to justify why
nodes as deep as these in the tree should unite taxa in a single genus, in
contrast to many other traditional genera with much shallower nodes. That the three are treated in 3 monotypic
genera signals that these are three old, highly divergent lineages.
B.
YES.
C.
YES.
D.
Option 1
E.
Option 1
F.
YES.
G.
Option 1. Option 5 (single genus) would
produce an exceptionally heterogeneous grouping the node for which is very deep
in the tree.”
Comments from Zimmer:
“A.
YES. From both vocal and
ecological (behavioral + habitat) perspectives, strigilatus, disjuncta
(especially). and pelzelni are each
unique and not clearly close to anything else.
I think that atrothorax is
most remarkable for its lack of unique characters, and hence, also does not
clearly align with any other member of this clade. Morphologically, atrothorax and pelzelni
are closest to one another in male and female plumages, but there just isn’t
that much there to justify lumping these disparate taxa into a single genus
that would essentially represent an unrecognizable hodgepodge.
B. YES
C. YES
D.
Option 1. YES. Option 2. NO, due
to the paraphyletic genus that would result if exsul is retained with the others, without also retaining griseiceps. Option 3. NO. I just think
that griseiceps is too different from the others, in virtually every
respect. Once we commit to keeping griseiceps in a monotypic Ampelornis, we’re forced into
recognizing Poliocrania and Sipia.
E. Option 1. YES. Option 2. No.
F. YES
G. Option 1. YES. This one presents the
toughest call for me. I would agree that
these three genera are somewhat weakly defined, although species-pairs within
each (e.g. arenarum-rufifrons, immaculata-zeledoni, goeldii-melanoceps) make intuitive sense
based on morphology, voice, and habitat/ecology. On the other hand, taken collectively (Option
3), they form a group that is also ill defined and somewhat squishy in my
opinion. Option 5 may be better (there
are lots of vocal and behavioral similarities that transcend current generic
lines and those proposed in Option 3), but I have always preferred recognizing
tighter, more internally cohesive genera to more inclusive, amorphous ones, and
I find it difficult to get rid of Gymnocichla
and the currently internally cohesive Pyriglena
(sensu stricto). While loudsongs of
nudiceps, immaculata and some others are suggestive of those of Pyriglena (as currently constituted), I
don’t think that any of the other species under discussion can approach leuconota (which, I’m pretty certain,
contains more than 1 good biological species), leucoptera and atra in
the complexity of their vocal repertoires, at least with respect to calls. These guys also seem to have more complex
social structures, as is usually the case within habitually ant-following
species. So, I think there is just too
much diversity to lump everything into a big Pyriglena, and I have similar objections to Option 3, which would
also be less internally cohesive than Option 5, due to similarities of Gymnocichla and Pyriglena to some members of what would be a diverse and expanded Percnostola. I think it comes down to either lumping
everything into one genus (Pyriglena),
which would be the most inclusive but also the most diverse and poorly defined
option, or, split into multiple genera as suggested by Isler et al. (2013), and
live with a bunch of small genera, a couple of which are weakly supported. Somewhat subjectively, I’m more comfortable
with the latter approach, especially given how the resulting genera would
relate to other long-recognized generic divisions within the Thamnophilidae.
Comments from Cadena:
“A. NO. These three taxa form a clade, so I think they are
best placed in a single genus. This arrangement would result in a
classification that is informative about phylogenetic relationships; that these
three species are closely related to each other is not something one can learn
from the classification if one places them in three monotypic genera. I realize
this situation might change in the future if any of the species turns out to be
split into more than one species (a reasonable expectation given our poor
understanding of species limits in antbirds), but we must act based on the
current situation. It was noted that lumping the three species in a single
genus would result in a heterogeneous entity relative to other genera in the
group. This point is well taken, but for the sake of consistency, are there any
other cases in our classification in which the only three species forming a
clade are each treated as members of monotypic genera?”
B. YES.”
Additional comments from
Robbins:“Given the
uncertainty of voting on 628G and Santiago’s recent vote, I revisited this
subproposal. As we all recognize this is
a subjective decision. Looking strictly
at the genetic data again, i.e., Fig. 1 from Isler et al (2013), I still favor
option 5 (and as I mentioned in my first round evaluation, because of the long
branch and unique plumage perhaps still maintaining Gymnocichla), but
option 3 would be my second choice.”
Additional comments from Remsen:
“G. Switching to Option 3, i.e.
included Akletos and Hafferia in Percnostola. I am persuaded by the comments above by those
in favor of Option 3.”