Proposal (753) to South
American Classification Committee:
Treat Poospiza whitii
as a separate species from P. nigrorufa
Effect
on South American CL:
This proposal would add Poospiza whitii to the SACC list, by splitting it from Poospiza nigrorufa.
Background: Three subspecies are
currently recognized in this complex: nigrorufa,
whitii, and wagneri, the latter of which has been considered doubtfully
distinct from whitii. Both nigrorufa and whitii were historically treated as full species until Hellmayr
(1938) lumped them by treating whitii as
the western subspecies of P. nigrorufa. Thereafter, nigrorufa and whitii were treated as a single geographically variable species but
by some authors as full species; see Jordan et al. (2017) for details.
Genetic data indicate that nigrorufa and whitii are well differentiated sister taxa with levels of mtDNA
divergence (ca. 2.5% for cyt-b and ND2) similar to those of other Poospizinae
sister species (Shultz and Burns 2013).
SACC proposal 79 failed to pass due to
lack of detailed published analyses, especially on vocal differences.
New
information:
Jordan et al. (2017) showed that
nigrorufa and whitii differ in 1)
plumage coloration and degree of dimorphism, 2) morphometric traits, 3)
habitat, distribution and ecological niche models, 4) vocal characters, and
that 5) in reciprocal playback experiments they ignore the other taxon while
answering strongly to their own vocalizations.
1) Plumage: In the slightly
dimorphic nigrorufa, males have tawny
rufous throat, breast and flanks, and are brownish grey with slate tinged upperparts (crown,
neck, back and rump), while females differ in the orange tinge of ventral parts
and in the more olivaceous upperparts. Sexes of nigrorufa are hard to distinguish both in field and museum
specimens. On the contrary, in the markedly dimorphic whitii, males have dark chestnut throat, breast and flanks, and
slate upperparts, while females exhibit tawny pale
orange throat, breast and flanks, and olivaceous
light-brown upper
parts (Fig. 1). In the field, females of whitii
exhibit paler ventral colors, and more olivaceous upperparts than both
sexes of nigrorufa. The key to
correct identification seems to be the extent of the white tip of the tail,
which is much greater in whitii than
in any sex of nigrorufa.
Jordan et al. (2017) also proposed that
the subspecies wagneri be treated as
a synonym of whitii because they were
unable to find any consistent plumage differences.
2) Morphometrics: Specimens of nigrorufa had significantly higher and
longer bill, longer tarsus and wings, and were~10% heavier than whitii (n=106 nigrorufa and n=91whitii).
Both species were sexually dimorphic, with males exhibiting longer wings and
tails than females. Within-sex comparisons between species show that nigrorufa has longer wings, but not
longer tails, than whitii (Fig. 2).
3) Habitat,
distribution and ecological niche models: Locality records (781 for nigrorufa and 322 for whitii) and models
of potential distribution indicate that ranges of both species are narrowly
allopatric, approaching closely in central Córdoba province (Argentina)
without overlapping (Fig. 3). P.
nigrorufa inhabits shrubby open areas in wetlands with reeds (Typha, Schoenioplectus) and bulrushes (Scirpus,
Rhynchospora) and grassy plains with
Pampas Grass (Cortaderia selloana),
whereas P. whitii inhabits closed to
semi-closed xerophytic to semi-humid scrub (Prosopis,
Geoffroea) and woodlands (Podocarpus, Alnus) far from wetlands. Finally, P. nigrorufa is generally a lowland species (except in southern
Brazil), whereas P. whitii is a
montane bird (Fig. 3).
4)
Vocalizations: Songs
(108 individuals; nigrorufa n= 81, whitii n= 27) and calls (18 individuals;
nigrorufa n=14, whitii n= 4) differed dramatically (Fig. 4). The simple song of nigrorufa consists of one phrase with
three pure whistled notes, that is repeated a variable number of times. This
phrase is usually transliterated as pleased
to meet you in English, quem te
vestiu in Portuguese, or bichi-bichi
in Spanish. The general shape and order of the three notes of the song of nigrorufa are very consistent throughout the range of the species.
The complex song of whitii is composed of a succession of a variable number of notes
whose quality and order varies greatly from individual to individual. In stark
contrast to nigrorufa, the number of
notes performed by each analyzed individual of whitii varied between 8 and 12. A complete song is a rhythmic
series of single melodious notes, among which paired melodious notes and a few
trills are irregularly interspersed that might be represented as choo we, tip-tip, sweet peer, tweak, trrree, sweet peer.
The numerous calls of both species are
easily distinguished with the aid of spectrograms, but are virtually impossible
to distinguish in the field (Fig 4.).
5) Reciprocal playback
experiments:
20 reciprocal sandwich-playback experiments were performed in Argentina during
the breeding season. In all experiments, the ten males of nigrorufa and the ten males of whitii
responded aggressively to conspecific vocalizations by approaching to the
sound source and singing, while ignoring heterospecific ones, regardless of
stimulus order (n=15 conspecific and 15 heterospecific stimuli for each
species).
Recommendation: We recommend a YES
vote. All lines of evidence clearly show that nigrorufa and whitii belong
to different species under any species concept. This long overdue split is now
fully justified with solid integrative evidence, including behavioral responses
to mating cues.

Fig.1.
A: P. nigrorufa male (Ph: Sebastián
Preisz) B: P. nigrorufa female (Ph:
D. Lins). C: P. whitii male (Ph: G.
Núñez), D: P. whitii female (J. La
Grotteria).
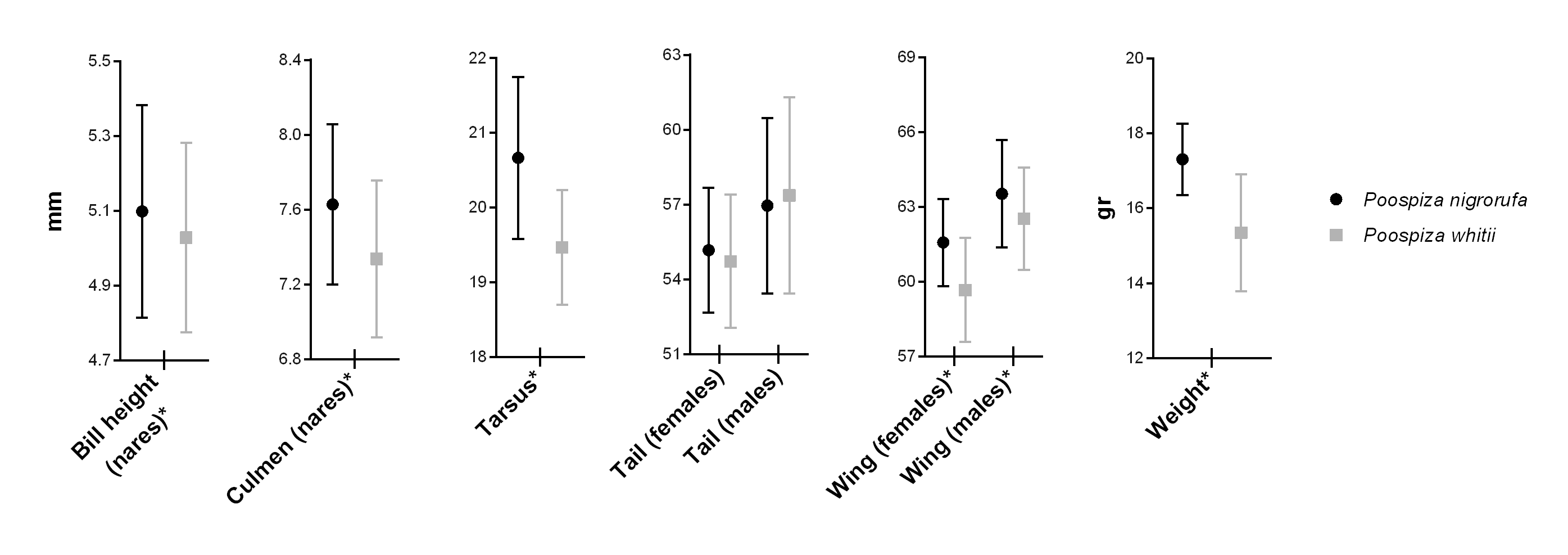
Fig. 2.
Morphological measurements and weight of Poospiza
nigrorufa and Poospiza whitii
showing mean and standard deviation. Asterisks denote significant differences
with alpha=0.05
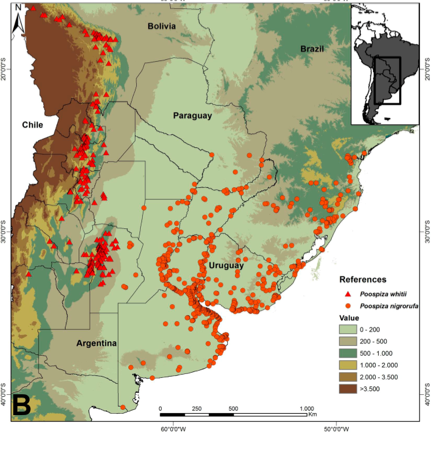
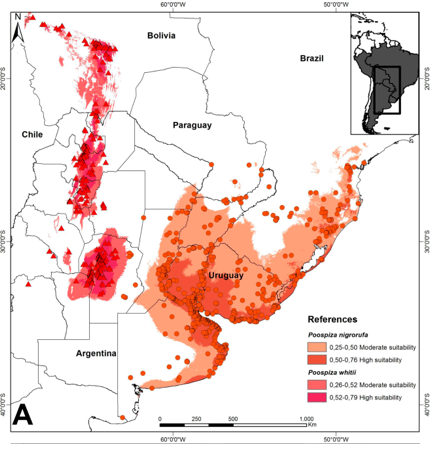
Fig.
3. Presence localities and potential distribution of Black-and-rufous
Warbling-Finch (Poospiza nigrorufa)
and Black-and-chestnut Warbling-Finch (Poospiza
whitii).
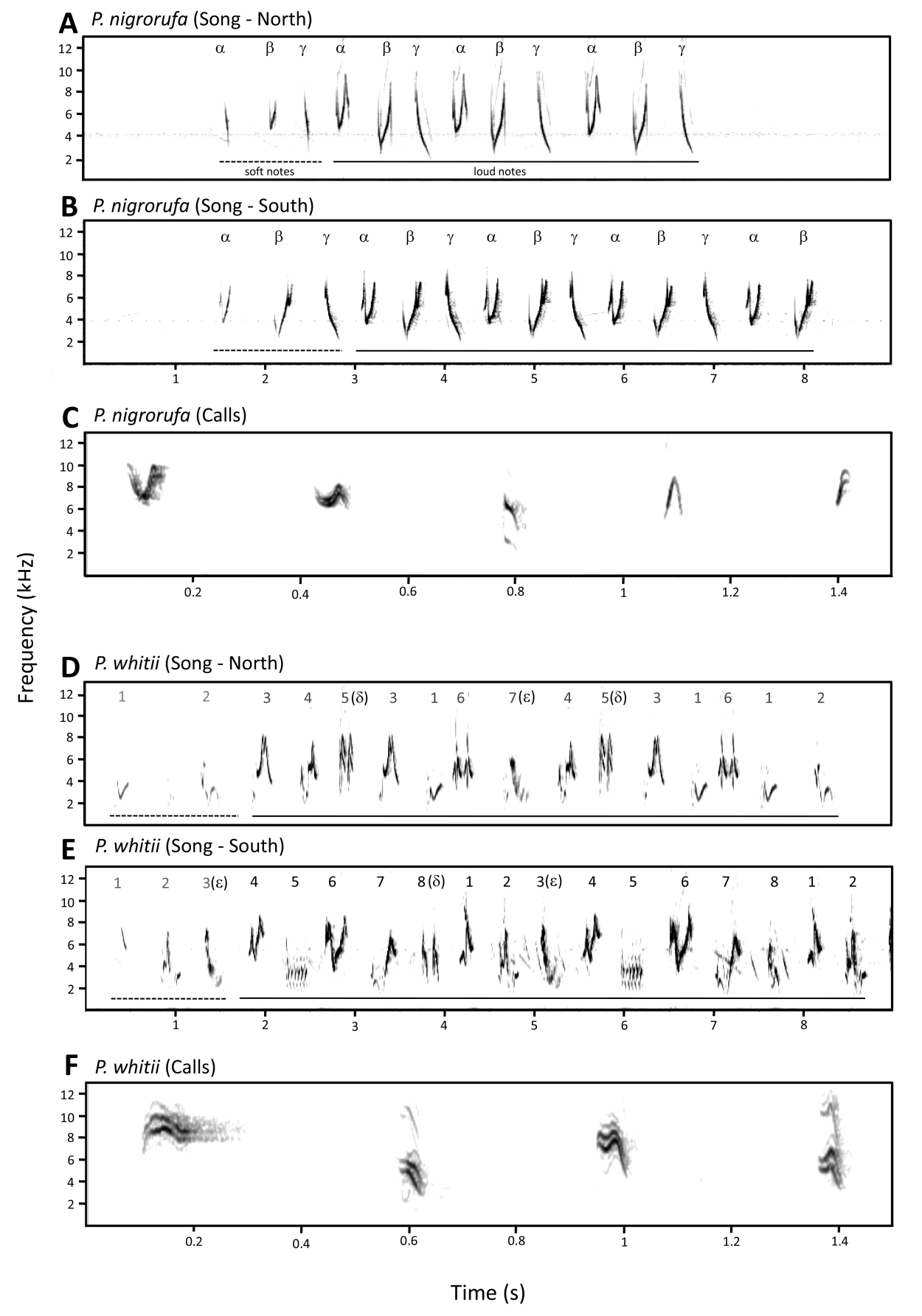
Fig. 4.
Spectrograms depicting songs (A, B, D, E) and calls (C, F) of adult males of
Black-and-rufous Warbling-Finch (Poospiza
nigrorufa) and Black-and-chestnut Warbling-Finch (Poospiza whitii). See Jordan et al. (2017) for further explanation
of symbols.
Literature cited
Hellmayr
(1938) Catalogue of birds of the Americas and adjacent islands. Field Museum of
Natural History Publications in Zoology 11, 1–662.
Shultz AJ
& Burns KJ (2013) Plumage evolution in relation
to light environment in a novel clade of Neotropical tanagers. Molecular
Phylogenetics and Evolution 66, 112–125. doi:10.1016/j.ympev.2012.09.011
Jordan EA,
Areta JI & Holzmann I (2017) Mate recognition systems and species limits in
a warbling-finch complex (Poospiza
nigrorufa/whitii). Emu
https://doi.org/10.1080/01584197.2017.1360746
Emilio
A. Jordan and Juan I. Areta
_________________________________________________________________________________________________________________________
Comments
from Stiles:
"YES. The combination of genetic, morphological and ecological data
clearly shift the burden of proof to those favoring conspecifity of whitii."
Comments from Zimmer:
“YES. Multiple data sets confirm the
distinctiveness of these two taxa, and the playback experiments confirm the
importance of the vocal differences, which were noted by Ridgely and Tudor
(citing R. Straneck) as far back as 1989.”
Comments from Remsen:
“YES. The playback trials in particular
are convincing evidence that these taxa have diverged to the point that free
gene flow no longer likely.”
“If this passes, then I suppose we should use
the names in Ridgely & Tudor that are already fairly entrenched and
included in the proposal above. However,
this is one of those cases in which the usual policy of creating new names for
both daughters should have received serious consideration.”
Comments
from Claramunt:
“YES.
The evidence is solid, in my opinion.”
Comments from Cadena: “YES. The work by Jordan et
al. is an an excellent, integrative taxonomic study. I do not think that
morphometric analyses revealing differences in measures of central tendency are
very useful for species delimitation (especially for allopatric populations)
and I think that ecological differences as revealed by niche models are
especially revealing in cases unlike the present one where potential
distributions suggest there is potential for populations to come in geographic
contact yet they remain distinct (we described this with Andrés Cuervo in a
paper in 2010 but the idea has not gained much traction). Nonetheless, all the
evidence points in the same direction that there are two different species
here.”
Comments
from Pacheco:
“YES. The evidence gathered is convincingly strong.”
Comments
from Robbins:
“YES, for recognizing Poospiza whitii as
a species based on multiple data sets.”