Proposal (754) to South
American Classification Committee:
Elevate 13 taxa to species rank based on playback
experiments
A. Elevate Pseudocolaptes
johnsoni to species rank
B. Elevate Automolus
virgatus to species rank
C. Elevate Grallaria alticola
to species rank
D. Elevate Scytalopus
intermedius to species rank [superseded by SACC 858]
E. Elevate Ochthoeca
thoracica to species rank
F. Elevate Myadestes
venezuelensis to species rank
G. Elevate Pheugopedius
schulenbergi to species rank
H. Elevate Amazonian populations of Tunchiornis ochraceiceps to species rank
I. Elevate South American populations of Basileuterus culicivorus to species rank
J. Elevate Myiothlypis
chlorophrys to species rank
K. Elevate Myiothlypis
striaticeps to species rank
L. Elevate Atlapetes
tricolor crassus to species rank
M. Elevate Amazonian populations of Arremon aurantiirostris to species rank
A prelude to a set of 13 proposals: background
and methods from Freeman & Montgomery (2017):
Background:
Geographically isolated populations of birds
often differ in song. Because birds often choose mates based on their song,
song differentiation between isolated populations constitutes a behavioral
barrier to reproduction. If this barrier is judged to be sufficiently strong,
then isolated populations with divergent songs may merit classification as
distinct species under the biological species concept.
Methods:
We conducted playback experiments to measure
whether populations discriminated against song from a related, allopatric
population - these experiments that simulated secondary contact between
geographically isolated populations. Briefly, each experiment measured the behavioral
response of a territorial bird to two treatments: 1) song from the local
population (sympatric treatment) and 2) song from the allopatric population
(allopatric treatment). All territorial birds responded to sympatric song by
approaching the speaker (typically to within 5 m).
We defined song discrimination as instances in
which the territory owner(s) ignored allopatric song, defined as a failure to
approach within 15 m of the speaker in response to the allopatric treatment. We
calculated song discrimination for each taxon pair as the percentage of
territories that failed to approach the speaker in response to allopatric song.
For example, a song discrimination score of 0.8 indicates that 80% of
territorial birds (e.g. 8 out of 10) ignored allopatric song while
simultaneously actively defending a territory. We assume that song
discrimination is a proxy for premating reproductive isolation; that is, our
experiments provide insight into whether these populations would recognize each
other as conspecific and interbreed (or not) were they to come into contact
with one another. It is unknown what degree of song discrimination is “enough”
that song constitutes a strong enough premating barrier to reproduction that
allopatric populations merit classification as distinct biological species. To
provide a yardstick, we considered nine allopatric Neotropical taxon pairs that
were recently split (or have pending proposals to SACC) in part based off
differences in vocalizations. We found the average song discrimination in these
nine taxon pairs to be ~ 0.6 (60% of territorial birds ignored allopatric
song), and suggest that species limits deserve to be reconsidered when taxon
pairs currently classified as subspecies have song discrimination scores above
~ 0.6.
In most cases, we played songs of populations A
and B (where populations A and B comprise a taxon pair) to territorial birds of
population A. That is, in the majority of cases we asked whether population A
discriminated against song from population B but not the reverse. To date, we
have conducted reciprocal playback experiments for 23 taxon pairs (13 oscines
and 10 suboscines, see Table S2) in which we measured both discrimination of
population A to song from population B and also discrimination of population B to
song from population A and in which we conducted at least four experiments on
each population (n = 11.5 ± 3.8 playback experiments/population, range = 4 – 23, see
Table S2). Song discrimination in these reciprocal cases was highly correlated
(r = 0.88, t = 8.4, df = 21, p-value
< 0.0001, Figure 1). There were no examples with strongly asymmetric song
discrimination (e.g. population A discriminates against population B, but
population B does not discriminate against population A). We therefore assume
that unidirectional data accurately describes song discrimination within taxon
pairs in our database.
Please forgive the brevity of the following
proposals. I did not have time to give each proposal the detail it deserves;
nonetheless, I believe the “bare-bones” approach of these proposals provides
sufficient relevant information to consider re-evaluating species limits in the
following 14 cases.
Benjamin
Freeman, 15 September 2018
754A.
Elevate Pseudocolaptes johnsoni to
species rank
Effect
on SACC:
Elevate Pseudocolaptes johnsoni to
species rank (split from Central American Pseudocolaptes
lawrencii).
Background: Related populations
of Pseudocolaptes (tuftedcheeks)
occur in Central America (lawrencii)
and western Colombia and Ecuador (johnsoni).
These allopatric populations are currently classified as conspecific, but
differ in plumage and voice. Due to these differences in mate choice traits,
they have sometimes been considered to represent two distinct species (e.g.,
HBW).
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 10 territories
of Central American lawrencii. They
found that 9 out of 10 territorial birds discriminated against song playback of
South American johnsoni
(discrimination = fail to approach within 15 m of the speaker. For canopy
species such as Pseudocolaptes, we
calculated distance to speaker in the horizontal plane. That is, a bird perched
high in the canopy but directly above the speaker would have a distance to
speaker of 0 m). Perhaps relevant is that these two populations differ in ND2
sequences (mtDNA) deposited on GenBank by 8.5% in uncorrected p-distance, which
suggests these two populations may have evolved strong postmating isolation as
well, although this is speculative.
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Pseudocolaptes johnsoni to
species rank (split from Central American Pseudocolaptes
lawrencii).
Recommendation: Central American
birds respond strongly to lawrencii
song, but essentially ignore song from johnsoni.
This suggests that vocal differences constitute a strong premating barrier to
reproduction between these taxa. I therefore recommend treating johnsoni and lawrencii as distinct biological species. English names currently
in use are Pacific Tuftedcheek for johnsoni
and Buffy Tuftedcheek for lawrencii.
However, at present “Buffy Tuftedcheek” refers to johnsoni + lawrencii. I
am agnostic on English names for this (and every other) proposal.
Additional
material added by Peter Boesman:
Morphological
differences were summarized in del Hoyo & Collar (2016): “Until recently, normally considered conspecific with P. lawrencii,
but differs in its stronger rufous (less buff-tinged) mantle and back (ns[1]);
rich rufous vs grey-streaked cream-buff underparts (3); rufous-tan vs
grey-black outer vanes of primaries (2); wing-coverts dark grey-brown with
vague rufous edges vs blackish with strong rufous tips (2); shorter tail
(sample size too small, but evidence indicative; ns).”
Vocal differences were treated by Spencer (2011) and Boesman (2016), and summarized in del Hoyo & Collar (2017): “divergent
song, being a high-pitched (score 2) rattled series of notes, slowing into
stuttering and ending (always) with a characteristic high-pitched down-slurred
note (2), vs several well-spaced staccato notes followed by a trill, which
usually ascends in pitch and then descends while slowing in pace (1).”
Playback
response was evaluated in one direction. Both Spencer
(2011) and (more systematically) Freeman & Montgomery (2017) found no response
of lawrencii to playback other than its own taxon-specific voice.
Comparative genetic differences: data is available for the three groups GenBank, apparently a genetic
tree which includes the three groups has not been published yet.
See:
Boesman, P. (2016). Notes on the vocalizations of
Buffy Tuftedcheek (Pseudocolaptes lawrencii). HBW Alive Ornithological
Note 87. In: Handbook of the Birds of the World Alive. Lynx
Edicions, Barcelona. https://doi.org/10.2173/bow-on.100087 https://static.birdsoftheworld.org/on87_buffy_tuftedcheek.pdf
Spencer, A. (2011). Variation in Tuftedcheek
vocalizations. https://www.xeno-canto.org/article/99
754B. Elevate Automolus virgatus to species rank
Effect
on SACC:
Elevate Automolus virgatus to species
rank (split from Amazonian Automolus
subulatus).
Background: Related populations
of Automolus woodhaunters occur west
of the Andes (virgatus group in
Central America and western Colombia and Ecuador), and east of the Andes (Amazon
basin; subulatus group). These
allopatric populations are currently classified as conspecific by SACC, but
differ markedly in voice, and somewhat in plumage. Due to these differences,
they have often been considered to represent two distinct species (e.g., HBW,
many field guides).
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 12 territories
of virgatus group individuals (6 in
Costa Rica and 6 in western Ecuador). They found that 11 out of 12 territorial
birds discriminated against song playback of the Amazonian subulatus group (discrimination = fail to approach within 15 m of
the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Split Automolus virgatus from A. subulatus.
Recommendation: Central American and
Choco birds respond strongly to local song, but essentially ignore song from
Amazonian birds. This suggests that vocal differences constitute a strong
premating barrier to reproduction between these taxa. I therefore recommend
treating the virgatus group and the subulatus group as distinct biological
species. English names currently in use are Western Woodhaunter for the virgatus group, and Eastern Woodhaunter
and Amazonian Woodhaunter for the subulatus
group.
Additional
material added by Peter Boesman:
Data available in 2003 were summarized in proposal https://www.museum.lsu.edu/~Remsen/SACCprop40.htm.
Besides the main remark that no peer-reviewed paper
had been published, there was also some uncertainty about cordobae. Since then, no in-depth study has been
published, but the following can be noted:
Morphological
differences were summarized in del Hoyo & Collar (2016) after studying a series
of specimens: virgatus group has duskier
underparts , with pale flammulations more confined to upper breast (1) and
slightly darker chestnut tail (1).
Vocal
differences have been qualitatively described in a variety of sources, were treated
more quantitatively in Boesman (2016), and summarized in del Hoyo & Collar
(2017): “virgatus group has a highly divergent song:
a series of evenly spaced identical short staccato notes
“keek..keek..keek..keek” vs a series of just a few (usually 2-3) downslurred
notes, often (perhaps when excited) followed by a low-pitched rattle, hence
more (ns 1) and much shorter notes (3) with a double peak frequency (2).”
Genetic differences have been documented in Claramunt et al. (2013). Although both groups
unsurprisingly are sisters (assimilis represents the virgatus group),
divergence seems to have occurred earlier than is the case e.g. in species of
the infuscatus complex.
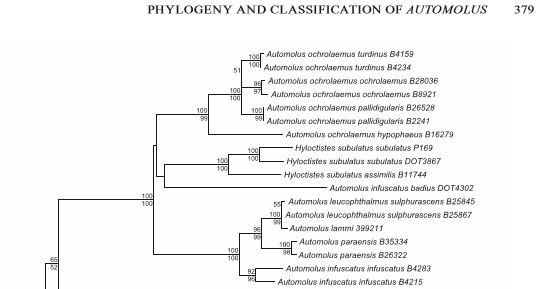
and more recently also documented by
Schultz et al. (2017):
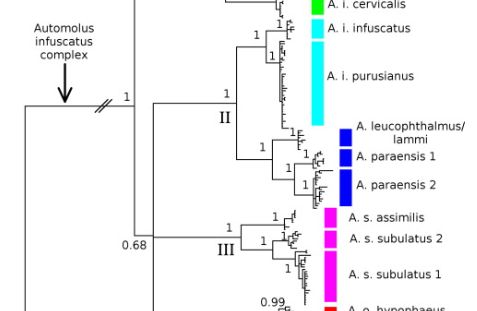
Although sonograms of cordobae now definitely
confirm it belongs to the western group, the remark in proposal 40 about a
peer-reviewed paper documenting voice remains. The incentive for such a paper
is, however. lower than ever. On the other hand, we have now a note quantifying
vocal differences, a paper documenting playback response, and genetic studies
indicating fairly high genetic divergence.
See:
Boesman, P. (2016). Notes
on the vocalizations of Striped Woodhaunter (Hyloctistes
subulatus).
HBW Alive Ornithological Note 90. In: Handbook of the Birds of the World Alive.
Lynx
Edicions, Barcelona. https://doi.org/10.2173/bow-on.100090 https://static.birdsoftheworld.org/on90_striped_woodhaunter.pdf
Claramunt,
S.; Derryberry, E.P.; Cadena, C.D.; Cuervo, A.M.; Sanín, S.; Brumfield, R.T.
(2013). Phylogeny and classification of Automolus foliage-gleaners
and allies (Furnariidae). Condor. 115 (2): 375–385.
Schultz,
E., Burney, C.; Brumfield, R.; Polo, E.; Cracraft, J., Ribas, C. (2017)
Systematics and biogeography of the Automolus infuscatus complex (Aves;
Furnariidae): Cryptic diversity reveals western Amazonia as the origin of a
transcontinental radiation. Molecular Phylogenetics and Evolution 107:503-515.
754C.
Elevate Grallaria alticola to species
rank
Effect
on SACC: Elevate Grallaria alticola to
species rank (split from G. quitensis).
Background:
Grallaria quitensis inhabits high
elevations (typically paramos) in the Northern and Central Andes. Three
allopatric subspecies differ in vocalizations, and, to a lesser extent,
plumage.
New
data: Freeman and Montgomery (2017) conducted playback experiments on 9
territories of quitensis in Ecuador
to measure response to alticola (restricted
to the Eastern Andes in Colombia). They found that 6 out of 9 territorial birds
in Ecuador discriminated against song playback of alticola (discrimination = fail to approach within 15 m of the
speaker). Perhaps relevant is that these two populations differ in ND2
sequences deposited on GenBank by 5.4% in uncorrected p-distance, which
suggests that the two populations may have evolved reasonably strong postmating
isolation as well, though this is speculative.
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Grallaria alticola to
species rank.
Recommendation:
Birds from Ecuador respond strongly to local song, but, in the majority of
cases, ignore song from andicola
birds. This suggests that vocal differences constitute a strong premating
barrier to reproduction between these two taxa. I therefore recommend treating alticola and quitensis as distinct biological species. I am not aware of
existing English names; HBW uses “Northern Tawny Antpitta” for alticola and “Western Tawny Antpitta”
for quitensis; there may be better
options.
Additional material added by Peter Boesman:
Morphological
differences between the 3 races were summarized
in del Hoyo & Collar (2016) after studying a series of specimens.
Vocal differences have been quantified in Boesman (2016) and summarized in del
Hoyo & Collar (2017). The two main
vocalisations (‘song’ and ‘call’) differ significantly between all 3 races.
They are also described in Greeney (2018).
Genetic data can be found for 2 races in Winger et al. (2015) and suggest paraphyly:
“Grallaria quitensis quitensis was identified as the closest out-group
to the bay-backed antpittas. However, the sample of G. quitensis alticola
was recovered as sister to G. hypoleuca (Fig. 2A), rather than to G.
q. quitensis, with low support”.
This case obviously reminds me of the
recent publications about the Rufous Antpitta complex, with the three races of G.
quitensis actually being the higher altitude replacement of resp. G.(r.)
rufula, G. (r.) saturata, and G. (r.) gravesi, and thus -even
more than the latter trio- separated by regions of unsuitable habitat.
We have here a very analogous
situation of limited morphological divergence and significant vocal differences
among allopatric populations separated by geographical barriers.
It boils down to the question whether a peer-reviewed
paper is considered a requirement or if sufficient analogy with a
well-documented case can be used as a convincing argument.
See:
Boesman,
P. (2016). Notes on the vocalizations of Tawny Antpitta (Grallaria quitensis).
HBW Alive Ornithological Note 72. In: Handbook of the Birds of the World Alive.
Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100072 https://static.birdsoftheworld.org/on72_tawny_antpitta.pdf
Greeney,
H. (2018) Antpittas and Gnateaters. Helm Identification Guides.
Winger
et al. (2015.) Inferring speciation history in the Andes with
reduced-representation sequence data: An example in the bay-backed antpittas
(Aves; Grallariidae; Grallaria hypoleuca s. l.) Molecular Ecology 24:6256-6277.
754D.
Elevate Scytalopus intermedius to
species rank
NOTE
from Remsen:
This proposal was superseded by SACC proposal 858.
Effect
on SACC:
Elevate Scytalopus intermedius to
species rank (split from S. latrans).
Background: Scytalopus latrans inhabits high elevations in the Northern and
Central Andes. Song is somewhat divergent between multiple populations. This
proposal focuses on two groups; the population of latrans found in southeastern Ecuador, and intermedius, which occurs in Peru south of the Marañon Gap.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 8 territories
of latrans in southeastern Ecuador.
They found that all 8 territorial birds in Ecuador discriminated against song
playback of intermedius (discrimination=
fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO=
Retain the status quo.
2)
YES
= Elevate Scytalopus intermedius to
species rank.
Recommendation: Birds from
southeastern Ecuador respond strongly to local song, but, in all cases, ignore
song from intermedius. This suggests
that vocal differences constitute a strong premating barrier to reproduction
between these two taxa, which are found on adjacent and opposite sides of the Marañon Gap. I therefore recommend treating
populations north and south of the Marañon Gap as distinct biological species. There
is marked variation in vocalizations within latrans
found north of the Marañon Gap, some of
which may be relevant in the future to species limits within the latrans group (= all population found
north of the Marañon Gap). I am not aware of
existing English name.
754E.
Elevate Ochthoeca thoracica to
species rank
Effect
on SACC:
Elevate Ochthoeca thoracica to
species rank (split from O.
cinnamomeiventris).
Background: Ochthoeca cinnamomeiventris is widely distributed in the tropical
Andes. Vocal and plumage variation is divergent between multiple populations.
This proposal focuses on two taxa; cinnamomeiventris,
found north of the Marañon Gap, and thoracica, which occurs in Peru south of
the Marañon Gap. These taxa differ in song and
plumage.
New
data: Freeman and Montgomery (2017) conducted playback experiments on 5
territories of cinnamomeiventris in
southeastern Ecuador. They found that all 5 territorial birds in Ecuador
discriminated against song playback of thoracica
(discrimination= fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Ochthoeca thoracica to
species rank.
Recommendation: Birds from
southeastern Ecuador respond strongly to local song, but, in all cases, ignore
song from thoracica. This suggests
that vocal differences constitute a strong premating barrier to reproduction
between these two taxa, which are found on adjacent and opposite sides of the Marañon Gap. I therefore recommend treating cinnamomeiventris and thoracica as distinct biological
species. An English name currently used for thoracica
is “Maroon-belted Chat-Tyrant”; as far as I can tell, cinnamomeiventris is known as “Slaty-backed Chat-Tyrant,” which is
also the English name widely used for the entire complex.
Additional
material added by Peter Boesman:
Vocal differences have been quantified in Boesman (2016) and summarized in del Hoyo &
Collar (2017).
(The case of nigrita remains to be solved
(treated as a full species by IOC). However, a recording of mine of an unseen
bird (XC430652) sounds very much like cinnamomeiventris, and seems to
indicate limited vocal divergence.)
Genetic data can be found in Garcia-Moreno et al. (1998) (in which they propose a
split): “The genetic differentiation between cinnamomeiventris and thoracica
(0.053) is of the same magnitude as that between other sister species (e.g.,
0.039- 0.063 within the S. diadema group, 0.042 between 0. leucophrys
and 0. oenanthoides”. Also Cuervo (2013) reached similar conclusions.
We have thus here a suboscine case of two groups which
differ moderately in morphology, significantly in voice, and show a fairly deep
genetic divergence.
See:
Cuervo,
A. (2013). Evolutionary Assembly of the Neotropical Montane Avifauna. Thesis,
Louisiana State University.
García-Moreno
J., Arctander P., Fjeldså J. (1998) Pre-Pleistocene differentiation among
chat-tyrants. Condor 100:629-640.
Boesman,
P. (2016). Notes on the vocalizations of Slaty-backed Chat-tyrant (Ochthoeca
cinnamomeiventris). HBW Alive Ornithological Note 253. In: Handbook of the
Birds of the World Alive. Lynx Edicions,
Barcelona. https://doi.org/10.2173/bow-on.100253 https://static.birdsoftheworld.org/on253_slaty-backed_chat-tyrant.pdf
754F.
Elevate Myadestes venezuelensis to
species rank
Effect
on SACC:
Elevate Myadestes venezuelensis to
species rank (split from M. ralloides).
Background: Myadestes ralloides is widely distributed in the tropical Andes.
Vocal variation is markedly divergent among populations of M. ralloides. This proposal focuses on two taxa; venezuelensis, found north of the Marañon Gap in eastern Ecuador, and ralloides, which occurs in Peru south of
the Marañon Gap.
New
data: Freeman and Montgomery (2017) conducted playback experiments on 12
territories of venezuelensis in
eastern Ecuador. They found that 10 out of 12 territorial birds in Ecuador
discriminated against song playback of ralloides
(discrimination= fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Myadestes venezuelensis to
species rank.
Recommendation: Birds from eastern
Ecuador respond strongly to local song, but, in nearly all cases, ignore song
from ralloides. This suggests that
vocal differences constitute a strong premating barrier to reproduction between
these two taxa, which are found on adjacent and opposite sides of the Marañon Gap. I therefore recommend treating
populations north and south of the Marañon Gap as distinct biological species. I am
not aware of existing English names. There is further vocal variation between
populations living on the Pacific slope in western Ecuador and those on the
Amazonian slope in eastern Ecuador – these may also prove to constitute
distinct biological species, but our (unpublished) playback experiments between
these taxa is not sufficient to evaluate species limits at present.
Additional
material added by Peter Boesman:
Morphological differences are analyzed in del Hoyo & Collar (2016) based on specimens in AMNH
and indicate a yellower bill and a paler belly, but evidently, differences are
rather small.
Vocal
differences have been quantified in Boesman (2016). Southern ralloides
appears vocally the most distinctive taxon, with plumbeiceps vs venezuelensis
also showing smaller vocal differences.
Genetic differences can be found in Miller (2007). “Average pairwise distances between M.
ralloides and M. coloratus (6.5%; range: 5.7–8.3%) and between M.
ralloides and M. melanops (6.0%; range: 5.9–8.0%) only slightly
exceeded the pairwise distance between M. ralloides populations across
the Marañon valley (6.0%; range: 5.3–7.3%). Pairwise distances between M.
coloratus and M. melanops averaged 2.0% (range: 1.4–3.6%), which is
similar to the shallowest divergence between phylogroups within M. ralloides.”
Cuervo (2013) reached similar conclusions.
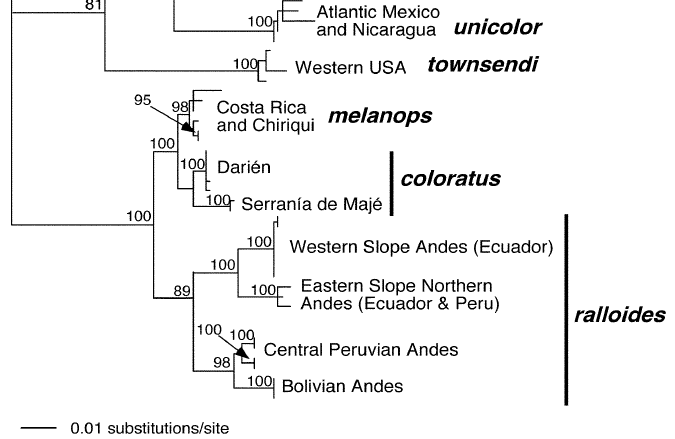
This is thus a case somewhat similar to Catharus
dryas in terms of differentiation (proposal 865).
See:
Boesman,
P. (2016). Notes on the vocalizations of Andean Solitaire (Myadestes
ralloides). HBW Alive Ornithological Note 435. In: Handbook of the Birds of
the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100435 https://static.birdsoftheworld.org/on435_andean_solitaire.pdf
Cuervo,
A. (2013). Evolutionary Assembly of the Neotropical Montane Avifauna. Thesis,
Louisiana State University.
Miller,
M., Bermingham, E., Ricklefs, R. (2007). Historical biogeography of the new
world solitaires (Myadestes spp.) Auk 124:868–885.
754G.
Elevate Pheugopedius schulenbergi to
species rank
Effect
on SACC:
Elevate Pheugopedius schulenbergi to
species rank (split from P. euophrys).
Background: Pheugopedius euophrys is widely distributed in the Northern Andes.
Vocal and plumage variation is somewhat divergent between multiple populations.
This proposal focuses on two taxa; longipes,
found north of the Marañon Gap in eastern
Ecuador, and schulenbergi, which
occurs in Peru south of the Marañon Gap.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 19 territories
of longipes in southeastern Ecuador.
They found that 12 out of 19 territorial birds in Ecuador discriminated against
song playback of schulenbergi (discrimination=
fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Pheugopedius schulenbergi to
species rank.
Recommendation: Birds from
southeastern Ecuador respond strongly to local song, but, in most cases, ignore
song from schulenbergi. This suggests
that vocal differences constitute a strong premating barrier to reproduction
between these two taxa, which are found on adjacent and opposite sides of the Marañon Gap. I therefore recommend treating
populations north and south of the Marañon Gap as distinct biological species. Schulenbergi is called “Grey-browed
Wren” by HBW.
Additional
material added by Peter Boesman:
Morphological differences are summarized in del Hoyo & Collar (2016).
Vocal
differences have been quantified in Boesman (2016).
Subspecies of P. euophrys show quite some
morphological differences in between them, as with schulenbergi. The
voice of schulenbergi is, however, remarkably different from all other subspecies,
and even for an oscine passerine with vocal learning ability this seems to be quite
a stretch to consider it a single species.
Some supportive genetic info is however highly
desirable.
There is also the case of the undescribed ‘Mantaro
Wren’, but until it is described as a new taxon, is better treated as a
potential future case.
See:
Boesman,
P. (2016). Notes on the vocalizations of Plain-tailed Wren (Thryothorus
euophrys). HBW Alive Ornithological Note 290. In: Handbook of the Birds of
the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100290 https://static.birdsoftheworld.org/on290_plain-tailed_wren.pdf
754H. Elevate Amazonian
populations of Tunchiornis ochraceiceps to
species rank
Effect
on SACC:
Elevate Amazonian populations of Tunchiornis
ochraceiceps to species rank (split from populations found west of Andes).
Background: Tunchiornis ochraceiceps consists of many named subspecies that are
found both west and east of the Andes. There is marked vocal and plumage
variation throughout its broad distribution.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 10 territories
of T. ochraceiceps in Costa Rica.
They found that 8 out of 10 territorial birds discriminated against song
playback from the western Amazonian ferrugineifrons
group (discrimination = fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Split populations found west of the Andes from populations found east of the
Andes.
Recommendation: Central American
birds respond strongly to local song, but essentially ignore song from western
Amazonian birds. This suggests that vocal differences constitute a strong
premating barrier to reproduction between these allopatric taxa. It would be
nice to have done further playback experiments in western Colombia and western
Ecuador, but this was not possible. There are additional vocal differences
within both west-of-the-Andes and east-of-the-Andes groups. In sum, though this
is not as clear-cut as many of the other cases, I recommend treating the
populations found east and west of the Andes as distinct biological species. I
am not aware of existing English names that would be applied to
west-of-the-Andes vs. east-of-the-Andes groups.
Additional
material added by Peter Boesman:
Vocal differences have been analyzed in Boesman (2016). Three vocal groups were
identified. NW group differs from Amazon group by having higher-pitched shorter
whistles. Guianan group differs from Amazon group by having two-note whistles
at different pitch.
Genetic data are given in Naka and Brumfield (2018).
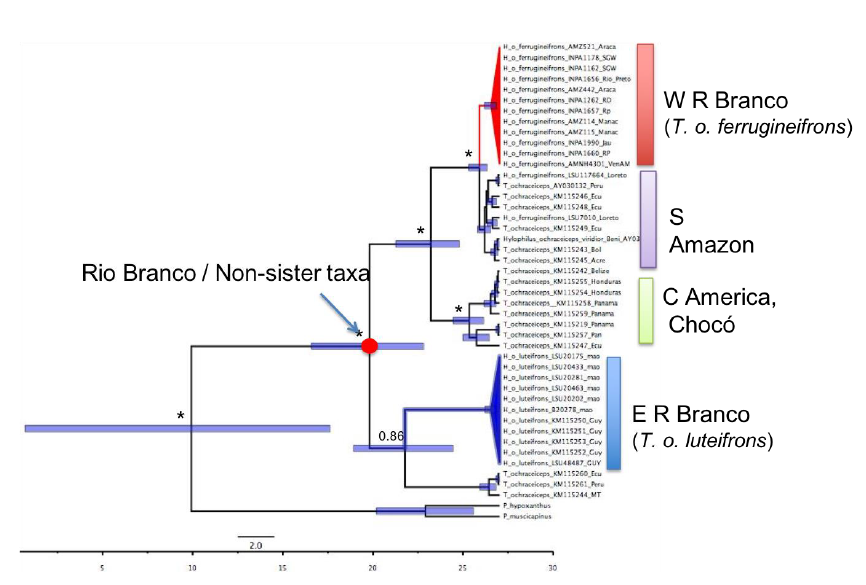
Genetic and vocal analysis seem to go hand-in-hand,
with luteifrons being most divergent, followed by NW populations vs
Amazon.
The situation of rubrifrons/lutescens is not
entirely clear however, and makes this case less clear-cut.
See:
Boesman,
P. (2016). Notes on the vocalizations of Tawny-crowned Greenlet (Hylophilus
ochraceiceps). HBW Alive Ornithological Note 168. In: Handbook of the Birds of
the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100168 https://static.birdsoftheworld.org/on168_tawny-crowned_greenlet.pdf
Naka,
L., Brumfield,R. (2018). The dual role of Amazonian rivers in the generation
and maintenance of avian diversity. Sci. Adv. 2018-4: 1-13.
754I. Elevate South
American populations of Basileuterus
culicivorus to species rank
Effect
on SACC: Elevate South American populations of Basileuterus culicivorus to species rank (split from populations
found in Central America).
Background:
Basileuterus culicivorus consists of
many named subspecies found throughout much of Central and South America. There
is marked vocal and plumage variation throughout its large distribution. This
proposal focuses on two allopatric taxa; godmani,
found in Costa Rica and Panama, and the cabanisi
group which occurs in nearby Colombia and Venezuela.
New
data: Freeman and Montgomery (2017) conducted playback experiments on 17
territories of godmani in Costa Rica.
They found that 13 out of 17 territorial birds discriminated against song
playback from the cabanisi group
(discrimination = fail to approach within 15 m of the speaker).
Competing
proposals:
1)
Retain
the status quo.
2)
Elevate
Basileuterus cabanisi to species
rank.
Recommendation:
Central American birds respond strongly to local song, but essentially ignore
song from cabanisi. This suggests
that vocal differences constitute a strong premating barrier to reproduction
between these allopatric taxa. Thus, I recommend treating the populations found
in South America as distinct from those in Central America. I note that species
limits within South American populations may require further evaluation.
English names: HBW uses “Stripe-crowned Warbler” for Central American
populations, “Yellow-crowned Warbler” for populations in Colombia and
Venezuela, and “Golden-crowned Warbler” for the additional South American
populations. We have no playback data to evaluate differences between the cabanisi group and other populations in
South America at this time. Still, the data we summarize above suggest that
Central American populations represent a distinct species from those in
adjacent northwestern South America; thus this proposal.
Additional
material added by Peter Boesman:
Vocal differences have been analysed in Boesman (2016). Three vocal groups were
identified. These seem to go
hand-in-hand with the three morphological groups (Curson et al. 1994),
and led del Hoyo and Collar (2016) to assign them species rank.
Genetic data is provided in Vilaca et al. (2010), who identified 5 clades.
Unfortunately, out of ca. 150 samples, only 2 were from the cabanisi
group (1 taxon). Nevertheless, it would seem that the South American subspecies
are a more recent colonisation from the East Mexico branch, which diverged
earlier from West Mexico. Thus, this seems to be a case in which evolution in
phenotypic traits have evolved at unequal rates in the distinct clades (which
as in other cases may lead to paraphyletic species).
(It is also suggested that B. hypoleucus may be
a colour morph of Golden-crowned Warbler, given there are no clear genetic of
vocal differences. This is however another case…)
See:
Boesman,
P. (2016). Notes on the vocalizations of Golden-crowned Warbler (Basileuterus
culicivorus). HBW Alive Ornithological Note 380. In: Handbook of the Birds
of the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100380 https://static.birdsoftheworld.org/on380_golden-crowned_warbler.pdf
Curson,
J., Quinn, D., Beadle, D. (1994). New World
Warblers. Helm Identification Guide. Helm. London.
Vilaca
S.T., Santos, F.R. (2010) Biogeographic history of the species complex Basileuterus
culicivorus (Aves, Parulidae) in the Neotropics. Molecular Phylogenetics
and Evolution 57: 585–597.
754J.
Elevate Myiothlypis chlorophrys to
species rank
Effect
on SACC:
Elevate Myiothlypis chlorophrys to
species rank (split from M. chrysogaster).
Background: Myiothlypis chrysogaster consists of two populations, one found in
the Choco in western Colombia and western Ecuador, and a second that inhabits
eastern Peru (and, narrowly, Bolivia).
These taxa are notably different in vocalizations and have been
considered to be distinct biological species by various authors.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 11 territories
of chlorophrys in western Ecuador.
They found that 10 out of 11 territorial birds discriminated against playback
of chrysogaster song (discrimination
= fail to approach within 15 m of the speaker). We also recently conducted two
playback experiments that showed that chrysogaster
discriminates against (ignores) chlorophrys
song (unpublished, and small sample size caveat, but I thought worth
mentioning).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Myiothlypis chlorophrys to
species rank.
Recommendation: Chlorophrys responds strongly to local song, but essentially
ignores song from chrysogaster. This
suggests that vocal differences constitute a strong premating barrier to
reproduction between these allopatric taxa. Thus, I recommend treating these
two populations as distinct biological species.
These taxa are sometimes referred to by the English names “Choco
Warbler” (for chlorophrys) and “Cuzco
Warbler” (for chrysogaster).
Additional
material added by Peter Boesman:
Proposal 68 (2003) already tackled this case. However, in those
days lack of sonograms etc. did not allow for a proper comparison.
Morphological differences were summarized in del Hoyo and Collar (2016): chlorophrys has a
broad green vs narrow yellow post-ocular supercilium, more olive flanks and
shorter tail.
Vocal differences have been analysed in Boesman (2016), identifying two very
distinct vocal groups (chrysogaster and chlorophrys).
Genetic data: Lovette et al (2010) only included the chlorophrys taxon. The two taxa may not have not been compared yet
genetically.
See:
Boesman,
P. (2016). Notes on the vocalizations of Golden-bellied Warbler (Basileuterus
chrysogaster). HBW Alive Ornithological Note 377. In: Handbook of the Birds
of the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100377 https://static.birdsoftheworld.org/on377_golden-bellied_warbler.pdf
Lovette
et al. (2010). A comprehensive
multilocus phylogeny for the wood-warblers and a revised classification of the
Parulidae (Aves). Molecular Phylogenetics and Evolution 57:753–770.
754K.
Elevate Myiothlypis striaticeps to
species rank
Effect
on SACC:
Elevate Myiothlypis striaticeps to
species rank (split from M. luteoviridis).
Background: Myiothlypis luteoviridis is widely distributed in the tropical
Andes. Vocal variation is somewhat divergent between multiple populations. This
proposal focuses on two taxa; luteoviridis,
found north of the Marañon Gap in eastern
Ecuador, and striaticeps, which
occurs in Peru south of the Marañon Gap.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 11 territories
of luteoviridis in southeastern
Ecuador. They found that 8 out of 11 territorial birds in Ecuador discriminated
against song playback of striaticeps (discrimination=
fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Myiothlypis striaticeps to
species rank.
Recommendation: Birds from
southeastern Ecuador respond strongly to local song, but, in most cases, ignore
song from striaticeps. This suggests
that vocal differences constitute a strong premating barrier to reproduction
between these two taxa, which are found on adjacent and opposite sides of the Marañon Gap. I therefore recommend treating
populations north and south of the Marañon Gap
as distinct biological species. I am not aware of existing English names.
Additional
material added by Peter Boesman:
Vocal differences have been analysed in Boesman (2016), identifying several vocal groups,
with striaticeps and euophrys especially standing out.
Genetic data: Lovette et al (2010) only included the euophrys taxon. Cuervo
(2013) provided a genetic tree in which the Marañon gap creates the earliest
separation of northern and southern groups (>3Mio yrs).
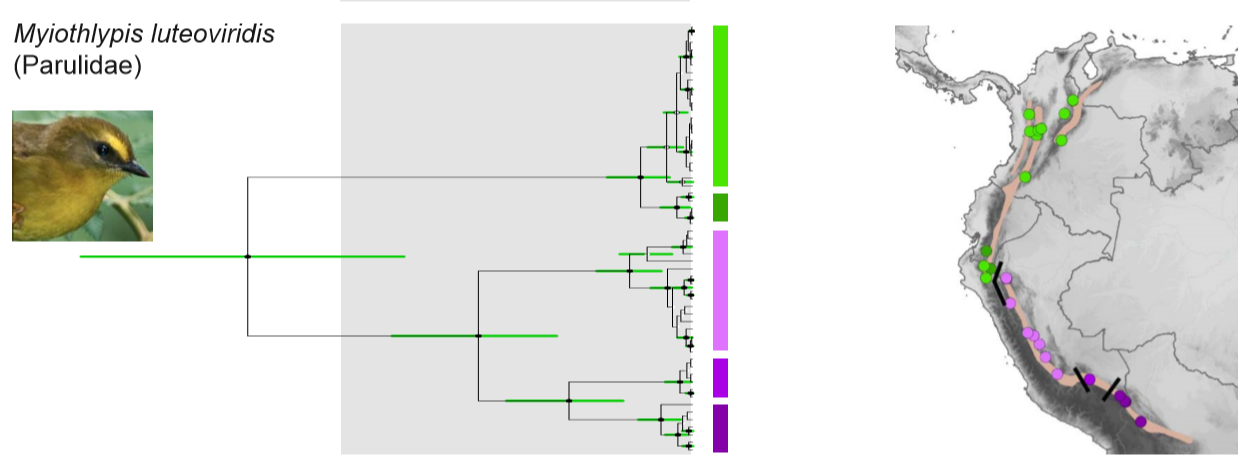
The southern subspecies striaticeps and euophrys
are thus clearly genetically and vocally distinct.
See:
Boesman,
P. (2016). Notes on the vocalizations of Citrine Warbler (Basileuterus
luteoviridis). HBW Alive Ornithological Note 438. In: Handbook of the Birds
of the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100438 https://static.birdsoftheworld.org/on438_citrine_warbler.pdf
Cuervo,
A. (2013). Evolutionary Assembly of the Neotropical Montane Avifauna. Thesis.
Lovette
et al. (2010). A comprehensive
multilocus phylogeny for the wood-warblers and a revised classification of the
Parulidae (Aves). Molecular Phylogenetics and Evolution 57:753–770.
754L.
Elevate Atlapetes tricolor crassus to
species rank
Effect on SACC: Elevate Atlapetes tricolor crassus to species
rank
Background: Atlapetes tricolor consists of two populations, one found in the
Choco in western Colombia and western Ecuador, and a second that inhabits
eastern Peru (and, narrowly, Bolivia).
These taxa are notably different in vocalizations (and morphology) and
have been considered distinct biological species by several authors.
New
data:
Freeman and Montgomery (2017) conducted playback experiments on 6 territories
of crassus in western Ecuador. They
found that 5 out of 6 territorial birds discriminated against playback of tricolor song (discrimination = fail to
approach within 15 m of the speaker).
Competing
proposals:
1)
NO=
Retain the status quo.
2)
YES
= Elevate Atlapetes crassus to
species rank.
Recommendation: Atlapetes crassus responds strongly to local song, but essentially
ignores song from tricolor. This
suggests that vocal differences constitute a strong premating barrier to
reproduction between these allopatric taxa. Thus, I recommend treating these
two populations as distinct biological species. “Choco Brush-Finch” has been
used for crassus and “Tricolored
Brush-Finch” for tricolor.
Additional
material added by Peter Boesman:
Morphological differences are summarized in del Hoyo and Collar (2016).
Vocal differences have been analysed in Sanchez-Gonzalez et al. (2015) and Boesman (2016).
Genetic data: Sanchez-Gonzalez et al. (2015) only included crassus,
and Klicka et al. (2014) only included tricolor. They were found at
different locations in the gene tree:
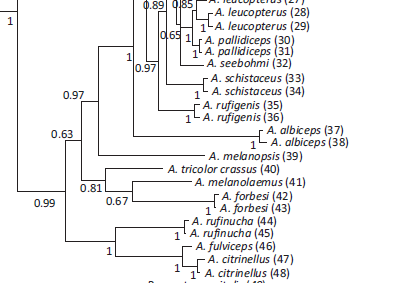
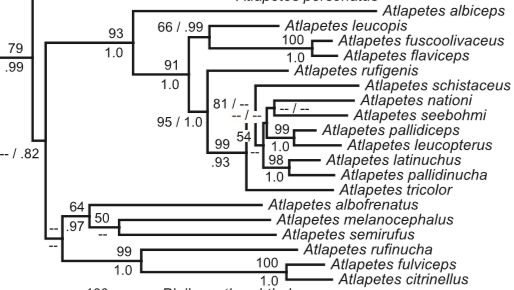
Sanchez-Gonzalez et al. (2015) summarizers: “In the
taxa investigated, the subspecies were found to differ little from each other
vocally, but one exception is A. t. crassus and tricolor, which
are distributed in western Colombia and Ecuador, and 900 km away in central
Peru. They also differ in elevational distributions 300–2000 m against
1750–3050 m) and morphologically (crassus with bill much larger, crown,
back and underparts differently coloured). We suggest that the northern form is
best treated as a full species: Choco Brush-Finch Atlapetes crassus. In
fact, the study by Klicka et al. (2014) included samples of A. t. tricolor
from Peru and recovered it as sister to our clade H, which includes Atlapetes
from a clade including mainly eastern Andean slope forms, widely separated from
our Ecuadorian A. t. crassus sample, thus confirming the taxonomic
differentiation in these two taxa.”
See:
Boesman,
P. (2016). Notes on the vocalizations of Tricolored Brush-finch (Atlapetes
tricolor). HBW Alive Ornithological Note 365. In: Handbook of the Birds of
the World Alive. Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100365 https://static.birdsoftheworld.org/on365_tricolored_brush-finch.pdf
Klicka,
J., Keith Barker, F., Burns, K. J., Lanyon, S. M., Lovette, I. J., Chaves, J.
A. & Bryson, R. W., Jr (2014). A comprehensive multilocus assessment of
sparrow (Aves: Passerellidae) relationships. Molecular Phylogenetics and
Evolution 77: 177–182.
Sánchez-González,
L.A., Navarro-Sigüenza, A.G., Krabbe, N.K., Fjeldså, J., García-Moreno, J.
(2015). Diversification in the Andes: the Atlapetes
brush-finches. Zool. Scripta 44(2): 135– 152.
754M. Elevate Amazonian
populations of Arremon aurantiirostris to
species rank
Effect
on SACC: Elevate Amazonian populations of Arremon
aurantiirostris to species rank (split from populations found west of
Andes).
Background:
Arremon aurantiirostris consists of
many named subspecies that are found both west and east of the Andes. There is
marked vocal variation throughout its distribution.
New
data: Freeman and Montgomery (2017) conducted playback experiments on 23
territories of occidentalis in
western Ecuador, and playback experiments on 12 territories of spectabilis in eastern Ecuador. They
found that 16 out of 23 territories of occidentalis
discriminated against playback of spectabilis
song, and that 8 out of 12 territories of spectabilis discriminated against playback of occidentalis song
(discrimination = fail to approach within 15 m of the speaker).
Competing
proposals:
1)
NO
= Retain the status quo.
2)
YES
= Elevate Amazonian Arremon aurantiirostris
spectabilis to species rank.
Recommendation:
Arremon aurantiirostris populations
in western and eastern Ecuador respond strongly to local song, but, in most
cases, ignore song from each other. This suggests that vocal differences
constitute a strong premating barrier to reproduction between occidentalis and spectabilis. Thus, I recommend treating these two populations as
distinct biological species. I note that this complex likely harbors additional
biological species (e.g., in Central America, populations on the Caribbean and
Pacific slope show strong song discrimination). Thus, if this split is adopted,
it should be recognized that this is not the end of the story (including
choosing English names that would minimize confusion in the case of additional
splits).
Comments
from Stiles: "This
is a most interesting proposal! However,
while highly suggestive of biological isolating mechanisms, in most of the
individual cases presented I consider that the data presented are insufficient
for SACC acceptance, for the following reasons:
1) Most of the cases presented involved
presenting the song of one population to members of another; reciprocal
playbacks would have been more convincing, as asymmetry in responses have been
found in some other studies;
2) The 60% no-response threshold presented was
insufficiently justified: just how much variation exists in this threshold
(which was based upon a general average of nine such comparisons of subspecies
vs. species responses); it would be helpful to know how much variation occurred
around this value, and whether the “threshold” might vary between oscines vs.
suboscines, especially given that several of the comparisons presented barely
exceed this “threshold”.
3) Several of the comparisons were made between
populations on either side of the Marañón gap; it is often not clear how close
these were to the “dividing line” (in at least one case, a subspecies closer to
the gap exists, but was not sampled). In other cases, the author did not
specify how closely the respective populations approach each other, and how
distant the sampled populations were from the point of closest approach;
4) No spectrographic analyses were made of
multiple recordings to evaluate possible intrapopulation variation, which might
influence the results in several cases; song dialects mediated by learning and
cultural transmission are known in oscines;
5) In several cases, important distributional
gaps occur between the populations sampled in which song variation is known to
exist between sampled and unsampled populations (subspecies), which could
easily influence interpretation of the results;
6) For nearly all cases, genetic data would
have helped to clinch the decision, but only for two cases were such data
presented;
7) The lack of nomenclatural precision often
makes the results difficult to apply, as in a number of cases, the taxa
involved are polytypic, which at a minimum would require a detailed study of
distributions and name priorities essential for arriving at concrete names for
the putative species; in several such cases, the subspecies sampled were not
given;
8) For a number of cases, pertinent
investigations are underway (or planned), especially where vocal differences of
unsampled populations may very well exist (a similar case in another SACC
proposal regarding Henicorhina
leucosticta did not pass for this
reason). I note that none of the experiments were performed in Colombia, which
would have been a strategic area for deciding several cases. Such cases in the
present proposal may best be taken as pointers for more comprehensive analyses.
"All
this is not to say that any of the cases presented are incorrect in suggesting
that two (or more!) biological species are involved, but the data are
insufficient for SACC action as they stand.
It is also worth noting that several of these proposals were rejected by
SACC due to the lack of detailed spectrographic analysis of vocalizations: the
point at issue is whether playback experiments represent a sufficient surrogate
or substitute for such analysis. I conclude that if the experiments show
significant differences in responses to same vs. different populations, the
results may be taken as valid. However, when the differences are very close to
the threshold value or sample sizes are too small, I recommend caution in their
acceptance. I now mention each case, with respect to the problems described
above by letters:
754A: Pseudocolaptes-plumage,
genetic and reciprocal playbacks all support this split, so YES.
754B: Automolus-data
very suggestive; not clear which population of the subulatus group was taken (where in “Amazonia”?). Were reciprocal playbacks done for the two virgatus populations, which are at
nearly opposite ends of this group’s range? On the other hand, in proposal 40
treating this question, all members of the committee agreed that two species
were indicated, given the vocal differences, but most preferred to await a
study of sonograms (by Zimmer?) Although I am not aware that this study has
been published, I consider that the experiments by Freeman constitute at least
a partial surrogate that to me, tips the burden of proof onto those who would
maintain these two as conspecific, hence YES.
754C: Grallaria-unfortunately,
no experiments performed in Colombia, where the Eastern Andes population also
differs vocally from that inhabiting the Central Andes, which would seem to be
in limbo: the same as the SE Ecuador population or not? Therefore, the data
here are incomplete; although genetic data suggest that the two populations
sampled represent different species, data from the C Andes birds in Colombia
are needed to complete the picture, and I suggest a tentative NO.
754D: Scytalopus-here
again, the data represent just the tip of the iceberg, since both north and south
of the Marañón gap there exist several subspecies with different plumages.
Daniel Cadena has been collecting genetic and vocal data on as many named Scytalopus taxa as possible, and may be
nearer to doing a comprehensive treatment of the latrans group. Hence, although the data show that at least two
biological species may well exist in this assemblage, I feel that any decision
should await more comprehensive data, so NO
for now.
754E: Ochthoeca-another
case where multiple, well-defined subspecies exist, especially N of the Marañón
gap, according to plumage; presumably the Ecuador sample was of subspecies; a
sample of only five experiments with thoracicus
from S of the Marañón and here again, I consider the data very suggestive
but incomplete: NO for now.
754F: Myadestes-nominate
ralloides was named from Bolivia
(from where were taken the recordings used for this form?); venezuelensis from Venezuela; SE Ecuador
is a long way from its type locality. Here, a detailed examination of
intraspecific variation in song from intermediate populations (e.g., in
Colombia and, depending on the above, N Peru) of these oscines (where song
learning and cultural transmission of song dialects could well occur) should be
done, and genetic data might also help: once again, I consider the experiments
presented here highly suggestive, but the data are as yet incomplete: NO for now.
754G: Pheugopedius-the
SE Ecuador population (longipes, N of
the Marañón gap) was tested against schulenbergi,
S of this gap: however, another subspecies (atriceps)
occurs closer to this gap than schulenbergi
and was not sampled. Intrapopulation variation in songs was not mentioned. So,
once more, the data are highly suggestive, but gaps remain. Also, the observed
non-response percentage (63%) was very close to the “threshold” (see above), so
NO for now.
754H: Tunchiornis-a
widespread species, the nominate subspecies found from Mexico to W Panama, whereas
cis-Andean ferrugineifrons occurs in
S Venezuela, Colombia and perhaps S to EC Peru. Although the exact site from
which the sample recording(s?) of the latter were taken was not given, evidence
for species status of cis- vs. trans-Andean populations sampled is certainly
suggestive. However, given the wide range of both forms, I am uneasy because
there could well be vocal variation within both forms; hence, I think that more
populations should be sampled for a conclusive answer: NO for now.
754I: Basileuterus
culicivorus-this is a highly polytypic species, with several named
subspecies in both Middle and South America. The tested population was godmani of Costa Rica to W Panamá, but
the provenance of the recording of the “cabanisi
group” was not specified (at least three subspecies of culicivorus occur in Colombia and Venezuela, and several more in
cis-Andean South America). Clearly, a much wider sample of the various
subspecies would be desirable, along with an evaluation of within-subspecies
variation in at least some of these. Population genomics within this species
would also be very interesting. Hence, I consider the available evidence too
incomplete to justify this split at present; NO for now.
754J: Basileuterus
chrysothlypis- This experiment dealt with a northern, trans-Andean
subspecies (chlorophrys) and a more
southern cis-Andean subspecies (nominate
chrysothlypis); these two are thus widely separated geographically, such
that parapatry is not relevant (or possible).
No other subspecies exist in this group. Here, the vocal evidence is
stronger (including a very small sample of reciprocal playbacks). Although
sonograms and genetic evidence would surely clinch the case, the available data
seem to me to tip the burden of proof towards conspecifity, hence a tentative YES.
754K:. Myiothlypis
luteiventris-Another comparison of subspecies N vs. S of the Marañón gap.
The song differences, as described by Schulenberg et al. (2009) certainly seem considerable, but with only 8 of 11
(73%) no-responses of the northern luteiventris
to songs of the southern striaticeps,
the results should be considered tentative at present. Also, a subspecies
further south (euophrys) shows
greater morphological differences from striaticeps
as well as possibly greater differences in song. So: NO for now, pending more data.
754L: Atlapetes-This case is similar to no. 10
in comparing cis- and trans-Andean populations, but with a much smaller sample
size (6); given that Atlapetes songs
tend to be complex, intraspecific variation could exist. I consider the
available evidence to be very suggestive but not conclusive, so NO for now.
754M: Arremon-This
is another tip-of-the iceberg case, because aurantiirostris
includes multiple subspecies with subtle morphological differences between
them over a wide range (including two from Costa Rica, that differ noticeably
in song). Hence, this result represents a small part of a much bigger problem.
Because Jorge Avendaño is now investigating this problem in depth, I believe
that any action by SACC should await the results of this study: NO for now.
Comments from Cadena: “I am going to vote NO on
all these proposals out of principle. I think that the hard work done by Ben
Freeman is exremely important and interesting, and is arguably the best
evidence we have to evaluate species limits in these groups of birds. However,
I agree with all the concerns voiced by Gary, especially with that related to
the 60% threshold for discrimination. I am not sure exactly what this means,
but say that 60% of the time, females of taxon A meeting males of taxon B
discriminate against them. What happens the rest of the time? If on 40% of the
encounters females of A do not discriminate against males of B, then inter-taxa
matings would be extremely frequent and one would expect hybridization to be
rampant, no? Maybe not, but then one should carefully consider other sources of
evidence together with the playback data, which takes me to a general
suggestion I have on all this. I think that the “bare-bones” approach taken by
Ben leaves several open questions and suffers importantly from lack of detail.
My suggestion is that all this should be placed in a paper (or papers) where
the ranges of the taxa involved are properly described, variation in different
characters is analyzed, and nuances of each case are discussed in detail to
reach more informed decisions. There are several potential theses projects here
for undergraduates with an interest in integrative taxonomy of birds in which
Freeman et al. have already made important progress.”
Comments from Jaramillo: “A – YES mainly due
to fact that this is a suboscine, and the genetic data.
B – YES
C – YES. The issue of
other populations in Colombia which may also differ, or perhaps muddy this picture
can be dealt with when those data appear.
D – YES. Scytalopus are so clear cut in response
to playback, that I find that this may be enough, even though there may be
other populations that should be sampled in the future. Or other molecular data
will arrive in the future.
E – NO. Kind of on the
fence on this one. But 5 experiments seems awful few.
F – YES. Note also that Miller et al. 2007 published
mtDNA data that also suggests a split north and south of the Marañon gap. Paper
is available here. https://www.researchgate.net/profile/Matthew_Miller3/publication/228644579_Historical_biogeography_of_the_new_world_solitaires_Myadestes_spp/links/00b7d521e0aefa5c7c000000.pdf
G – NO. I would like to
see more data here, 12 out of 19 is a lot of birds that did react positively.
More data is necessary in this case.
H – NO. Given the huge
variation in this species, I think a more widespread set of experiments, is
necessary.
I - NO, but not because I do not believe that
there are multiple species here. But this one is one of those complex ones with
possibly multiple species involved, furthermore B. hypoleucus may in fact be conspecific with one of these forms.
So, it is complicated enough that I would rather base changes on the Vilaca
& Santos (2010) paper. Copy available here: https://s3.amazonaws.com/academia.edu.documents/46692562/Biogeographic_history_of_the_species_com20160621-14843-6idjqj.pdf?AWSAccessKeyId=AKIAIWOWYYGZ2Y53UL3A&Expires=1522809006&Signature=X%2FzDXcNowKX2AzmZ6ykEM8dpuNc%3D&response-content-disposition=inline%3B%20filename%3DBiogeographic_history_of_the_species_com.pdf
J – YES
K – YES, but tentative.
L – YES. Available
molecular data corroborates. Sanchez-Gonzales et al. (2014) found that crassus was in a group with A. melanolaemus and A. forbesi. While Klicka et al. (2014) sampled A. tricolor tricolor and found it was in an entirely different
clade with schistaceus, latinuchus
etc. Sanchez-Gonzales et al. (2014) also confirm clearly defined differences in
song of tricolor and crassus.
Klicka, J., Keith
Barker, F., Burns, K. J., Lanyon, S. M., Lovette, I. J., Chaves, J. A. &
Bryson, R. W., Jr (2014). A comprehensive multilocus assessment of sparrow
(Aves: Passerellidae) relationships. Molecular Phylogenetics and Evolution, 77,
177–182.
M- NO. Playback
experiments not quite conclusive here, and a very complex situation.
Additional
comments from Stiles:
“I recommended a YES on a few (!) of these, because I feel that they did tip
the balance on already accepted but
still controversial splits (i.e., not accepted by SACC but widely implemented
elsewhere). I also don’t feel
comfortable with a rather dogmatic NO to all without consideration of the
contexts involved.”
Comments
from Areta:
“I am also going to vote NO on all proposals, and I agree
with Daniel’s views. I agree with all the concerns voiced by Gary, and while I
appreciate the effort spent by both Alvaro and Gary in analyzing each case
separately, these analyses are not as thorough and complete as I would like to
have before me to make an informed decision. I especially feel that the 60%
threshold is rather artificial and insufficient. For example, by adding data on
reciprocal playback experiments of Upucerthia dumetaria/saturatior, Pseudocolopteryx
citreola/flaviventris and Poospiza nigrorufa/whitii (all of which
show 100% discrimination) this value can change easily. Not to mention other
Suboscines in which responses (even between sister species) are all or none.
Also, it troubles me that there are no reports of cases in which the local
population ignored its own local song and no assessment of whether the
playbacks were done at comparative times of the year (e.g., peak breeding
season, etc.). In sum, all these playback experiments are informative to
understand some key behavioral traits of possibly separate species, but the
taxonomically focused works needed to put these results in perspective are
missing (i.e., assessment of validity and priority of names, wide geographic
sampling, understanding of other phenotypic traits, evaluation of vocal traits
of intervening and/or other closely related populations, possibly genetic data,
etc.).”
Comments from Claramunt: “I agree with the several caveats raised by Gary regarding the evidence
presented by Freeman & Montgomery. In particular, unless there is evidence
of homogeneity within taxa, I’m skeptical about playback experiments involving
only two populations that are far apart as a way of assessing reproductive
isolation. At least, populations in close proximity or most likely to be in
secondary contact should be evaluated. Only interpreted together with other
information, the results of Freeman & Montgomery help to tip the balance in
some cases, as Gary stated. I think we should take advantage of this
opportunity to make decisions regarding species limits in light of this new bit
of information. Waiting for the ideal playback experiments is not an option, in
my opinion.
“A. YES to elevate Pseudocolaptes johnsoni to
species rank. With a
darker and more rufous plumage and a distinctly decurved culmen, johnsoni could be considered the most distinctive
taxon in its genus. As confessed by Zimmer (1936), it was only because of its
rarity (and scarcity of specimens) plus the uncertainty regarding its type
locality what made him treat johnsoni as a subspecies of lawrencii,
despite believing that it was a good species. Taxonomic inertia followed (with
few exceptions). Now we know that the song of johnsoni is very different and is not recognized by lawrencii.
A necessary and long overdue split.
“B. YES to
elevate Automolus virgatus to species rank. This complex shows considerable phenotypic variation that deserves
thorough study. Birds from the Choco region (assimilis) are darker and do not have the distinctive slender beak
of subulatus; they have instead a
more Automolus-like beak. However,
the slender beak, similar to that of subulatus,
reappears in Central America in virgatus,
although birds look much darker. A new relevant paper should also be
considered: Schultz et al. 2017 (https://www.sciencedirect.com/science/article/pii/S1055790316304468). Schultz et al. 2017 found
that subulatus and assimilis represent two distinct genetic
groups divided by the Andes. Other subspecies were not sampled. mtDNA shows two
reciprocally monophyletic groups corresponding to subulatus and assimilis.
Nuclear data is harder to interpret because assimilis
was only sequenced for BF7, and BF7 alleles are messy across the entire genus.
However, the relevant information is that no BF7 alleles were shared between subulatus and assimilis (Supplemental Figure C7), thus suggesting no recent gene
flow. The clustering algorithm used (BAPS) failed to detect this separation
probably because of the missing nuclear data for the trans-Andean populations.
A pending question is whether assimilis
is indeed conspecific with virgatus
or represents a third species, an issue that could be addressed by studying the
Panamanian populations, where a contact zone could exist. In the meantime,
splitting Amazonian from trans-Andean forms is a step forward.
“C. NO to elevate Grallaria alticola to species rank. Playback
experiments show some degree of discrimination, but populations tested are far
apart. If song structure or song preference vary geographically, then the
experiments are irrelevant. No other traits differentiate Grallaria alticola,
except it body size, but a formal analysis of geographic variation is required
to determine if variation is continuous or discrete.
“D. NO to elevate Scytalopus intermedius to species rank.
Skimming over what is available in xeno-canto suggests tremendous and complex
variation in vocalizations in SE Ecuador and N Peru. The test based on just two
populations in this complex is not very informative of what is going on in the
region. Much more work is needed.
“E. YES to elevate Ochthoeca thoracica to species rank. Although
the number of independent experiments was low (5), this information should be
interpreted in light the additional information available. Plumage and song
variation is prominent and discrete, and matches the taxonomic divide. To the
eyes of the subspecies-lover ornithologist of the mid XX century, this
variation was only worth subspecific distinction. Freeman and Montgomery (2017)
showed that for the birds themselves, this variation means much more, and
suggest that these two separate lineages are reproductively isolated because of
divergent mating signals. Furthermore, a mitochondrial tree by Andres Cuervo (https://digitalcommons.lsu.edu/gradschool_dissertations/275/),
including a large sample across the entire distribution of this complex, shows
that thoracica and cinnamomeiventris are reciprocally monophyletic, combining a
high degree of divergence and low intra-taxon genetic variability. Therefore,
multiple lines of evidence (phenotypic distinctness, playback experiments,
genetics) demonstrate that this pair of taxa constitute separate species.
Addendum. Note that Ochthoeca
thoracica and Ochthoeca cinnamomeiventris were treated as separate
species by Cory & Hellmayr and they were lumped later by Peters without
providing a rationale. So, this proposal should not be viewed as few playback
experiments in isolation but as evidence for whether two taxa that are
differentiated in plumage, song, and mtDNA are able to discriminate each other
based on playback experiments. Although sample size is not great, the answer
seems yes. So, for this case that has been historical in the borderline we
finally have evidence of behavioural reproductive isolation.”
“F. NO to elevate Myadestes venezuelensis to species rank.
The playback experiments are suggestive but geographic variation in other
traits needs to be studied.
“G. NO to elevate Pheugopedius schulenbergi to species
rank. Song discrimination was partial and geographic variation in songs and
plumage is high across the complex.
“H. NO to elevate
Amazonian populations of Tunchiornis
ochraceiceps to species rank. There is much geographic variation in this
complex and a test of reproductive isolation based on just two distant
populations is not very informative. Moreover, the mitochondrial tree of Naka
& Brumfield (2018, http://advances.sciencemag.org/content/4/8/eaar8575) indicates that Amazonian populations are paraphyletic because
trans-Andean populations are nested within them. Therefore, the situation is
complex and warrants more studies before redefining species limits.
“I. NO to elevate South
American populations of Basileuterus
culicivorus to species rank. Song discrimination was partial, and the
population tested was far away from the potential contact zone. Also,
geographic variation in songs and plumage is high across the complex, and a
phylogeographic study did not recovered the central American taxa as
monophyletic (Vilaça & Santos 2010, Mol. Phyl. Evol. 57:585-597).
“J. YES to elevate Myiothlypis chlorophrys to species rank.
I don’t have much additional information on this case. The two taxa are widely
allopatric. Plumage differences seem subtle, but that is typical in this genus.
On the other hand, songs differ dramatically and Freeman and Montgomery (2017)
showed that birds discriminate strongly against songs from the other taxon.
“K. NO to elevate Myiothlypis striaticeps to species rank.
These forms are phenotypically very similar, and some Ecuadorian birds did
respond to alien songs. So, I don’t see compelling evidence for a separation.
“L. YES to elevate Atlapetes tricolor crassus to species
rank. Not mentioned by the proposal is the possibility that tricolor and crassus are not each other’s closest relatives: tricolor seems to be related to schistaceus, seebohmi and related species (Klicka et al. 2014 Mol. Phyl. Evol. 77:177-182), whereas crassus forms a clade with melanolaemus and forbesi (Sánchez González et al. 2015 Zoologica Scripta
44:135–152). I know that this does not make sense biogeographically or plumage-wise,
and needs to be corroborated by an analysis in which the two forms are analyzed
together. But taken together, songs, phylogeny, and playback experiments all
point to separate status for these two forms despite plumage similarity, which
we know is highly homoplastic in Atlapetes.
“M. NO for now to
elevate Amazonian populations of Arremon
aurantiirostris to species rank. We have a small series of specimens of at
the ROM, and it strikes me as very distinct: short and decurved culmen, orange
underwings (instead of yellow), and other details. I think that it is a
different species. Not sure how it was decided that it belongs into aurantiirostris. However, it has been
treated as a subspecies of aurantiirostris
forever. and playback experiments show that they recognize each other to some
degree, so I would wait for the forthcoming more detailed study.”
Summary
comments from Peter Boesman (see individual contributions to each subproposal
in the main proposal):
“I personally believe that -staying at the conservative side- at
least case A, B, E, F, J and L should go through. (I am also quite convinced about C and G, but
am aware there is little published evidence)”
Additional
comments from Claramunt of 754C: “Regarding Grallaria
alticola, a clarification regarding the phylogenetic evidence: only in the
mtDNA the two samples analyzed by Winger et al. (one of alticola and one
of quitensis) did not appear as sister, but this results lacked strong
statistical support. The genomic dataset, in contrast, showed the two samples
as sister with moderate to strong support (Fig. 2B). Therefore, although the
genetic distance between the two samples was high in both cases, it seems that
they are sister taxa.
“Morphological differences seem slim, as far as I can see, and the
playback experiments revealed only partial discrimination, suggesting that the
differences in songs are not that significant for the birds.
“So, I maintain my NO on this sub proposal.”
New
comments from Jaramillo: “Very little change. No comments
on the ones where there was no change. Some additional comments where confusion
arose or perhaps now data is better but suggests a more complex re-organization
that may require a new proposal? Here we
go:
A – YES
B – YES
C – YES.
D – YES.
E – YES -
this is a change from my previous vote. Based on the new information,
characterization of vocal differences and published genetic data
F – YES
G – NO
H – NO. I
am confused here now. No change from my previous voting. However, it seems like
the new genetic and vocal data suggests that the real outlier is the Tepui
population. So perhaps this should be expanded as a new proposal? I am not sure
how to deal with this one. But something is going on here.
I - NO, see
my previous comments. May require re-writing this one as a new proposal? More
information needs to be unpacked for this one.
J – YES
K – YES,
now stronger based on new data.
L – YES.
M – NO.
Playback experiments not quite conclusive here, and a very complex situation.”
New comments from Stiles: “To
begin with, I believe that it is incumbent upon us to take this proposal
seriously enough to evaluate each subproposal based not only upon the playbacks
themselves, but such other evidence as exists in each case. Much additional evidence
has been gathered by Boesman for most of the proposals that merit
consideration. I am impressed by his careful evaluation of vocal differences in
particular, and he has brought to bear several pertinent sources not readily
available (e. g., Andrés Cuervo’s thesis). With reference to Santiago’s comment
that Freeman did not report results of playbacks of one population to its own
songs, Freeman stated that all such “sympatric” playbacks produced strong
positive reactions (approach to 15 m or less), although he did not give the
number of same for each case. Considering that the alternatives in each case
are to retain the status quo (a single species) or split this into two, I think
that one must decide whether the information presented is sufficient to tip the
burden of proof from those favoring the split onto those who would maintain the
status quo. However, this with the qualifier that although the evidence may
favor the two-way split, the proposal is embedded within a wider problem
requiring further work to disentangle. I might also mention that in some cases,
the method used for the playbacks could bias the results. A positive result for
an “allopatric” experiment (i.e., playing the voice of one population to a bird
of a distinct population) could produce some degree of approach simply because
of its novelty to the recipient: in effect, mere curiosity rather than an
aggressive response.
“On to specifics: my votes for all subproposals:
A.
YES to split Pseudocolaptes
johnsoni from P. lawrencii. Boesman
has marshalled morphological, vocal and genetic data to clinch this case.
B.
YES to split Automolus
virgatus from A. subulatus. Again,
Boesman presents a more convincing case.
C. NO,
albeit barely – burdens of proof nearly even on both sides. The vocal evidence
of Boesman for recognizing three
species in G. quitensis is quite
convincing, however genetic evidence is incomplete and inconsistent (no single
study has included all three, and sample sizes here are limited: denser
sampling accompanied by including nuclear genes and haplotype sharing would
certainly clinch the case, even given the subtle morphological differences.
Strongly suggestive of the G. rufula
study!
D. Already
done by Krabbe (proposal no. 858).
E.
YES to split Ochthoeca
thoracicus from O cinnamomeiventris –
additional evidence gathered by Boesman definitely tips the balance.
F.
NO, albeit tentatively, to splitting Myadestes venezuelensis from M. ralloides. I am on the fence here.
Genetic evidence favors the split, but could be bolstered by including nuclear genes
and haplotypes, and intraspecific variation in songs and calls should be
evaluated with larger sampling from all subspecies. Morphological differences might be given more thorough examination
as well.
G. NO
to splitting Pheugopedius schulenbergi from P. mystacalis, basically for the same
reasons: gaps in both vocal and genetic evidence that should be filled in to
clinch the case.
H. NO
to splitting Tunchiornis into cis-
and trans-Andean species, at least until the biogeography and intraspecific
variation can be sorted out more completely. The genetic evidence suggests that
three or four species could be recognized, in line with vocal evidence briefly
mentioned by Boesman, but the biogeography looks complicated. Claramunt noted
that the Mesoamerican-Chocó population appeared to be genetically embedded
within the two “Amazonian” populations, but this seems to me to be a function
of how the branches are ordered: flip Centromerica-Chocó to the top, (above
that including the W of rio Branco and south Amazon) and you get a
north-to-south ordering of these three, which seems biogeographically logical.
The fly in all this ointment is the Guiana (E of rio Branco) population: it is
widely divergent from the population the other side of the river, as well as
that of Centroamerica-Chocó, so how and when did it reach its present range
without passing through the area occupied by the latter? If the tree given by
Boesman was exclusively for mitochondrial genes, adding nuclear genes and an
analysis of haplotype sharing might clarify the picture.
I.
NO to splitting Basileuterus
culicivorus into 2?3? species. The vocal analysis by Boesman is suggestive,
but taxon sampling could be improved; the genetic data for South America in
particular are decidedly sparse; especially lacking are data from Colombia,
where the two populations (godmani
and cabanisi) may come into contact.
J.
YES to split Myiothlypis
chlorophrys from M. chrysogaster. Boesman
demonstrates pronounced vocal differences; the morphological differences are
roughly on a par with those between several other species pairs in this genus;
and the wide latitudinal separation on different slopes of the Andes would be
most unlikely for a single species.
K.
A tentative YES to splitting Myiothlypis striaticeps from M.
luteoviridis. Morphological
differences exist (at least comparable to the previous case), and the vocal
analysis by Boesman suggests differences between these; especially notable are
the genetic data by Cuervo, which indicate a very deep split between these
lineages on opposite sides of the Marañón, which to me tips the balance of
proof.
L.
YES to splitting Atlapetes
crassus from A. tricolor:
detailed genetic data demonstrate that the two are not close relatives within Atlapetes, let alone sisters.
M. NO
to splitting E Costa Rican and E Ecuadorian populations of Arremon aurantiirostris into two species; here there are several
other subspecies to be considered; genetic and morphological data are sparse
and taxon sampling insufficient. (Unfortunate that while in Costa Rica, Freeman
did not do experiments between the Caribbean and Pacific populations of the
species, which have quite different songs.)”
Comments from Lane:
“ A. NO, although Boesman’s comments, in conjunction with the newly
published Harvey et al. phylogeny (which would suggest that the two taxa are
*not* sisters) would probably be enough to tip the scales if presented in a
revised proposal.
B. NO. The playback used sounds like it only compared trans-Andean birds
to playback of cis-Andean birds, and not with playback from other named
trans-Andean taxa, if I am reading the original paper correctly. This seems a
bit of an oversight, and would have been a useful datapoint to include.
C. NO. Again, there is an additional taxon, atuensis, which should
also be considered for species status.
E. NO. There are multiple taxa within this complex, and it is not clear
what to do with several of them following this split.
F. NO. Once again, there are several taxa in addition to the two
considered here, and where they would fit into the split is unclear
G. YES. I am actually inclined to consider this case as enough evidence
to separate the very differently voiced schulenbergi as a separate
species from the rest of the taxa presently under the name euophrys,
with the understanding that there may be further taxonomic assessment necessary
later for the remainder of the complex. I would say, in response to Boesman’s
comment on the Mantaro bird, that its inclusion with euophrys rather
than eisenmanni is speculative at best and, in my opinion, incorrect.
But, until that population is named, it can’t be considered as a player in this
game.
H. NO. The number of taxa involved makes this a messy case that cannot be
resolved with such limited.
I. NO. There are many taxa involved, both in Middle America and South
America, and this playback experiment barely scratches the surface.
J. YES. This is another case where I believe the differences are so stark
that this one-way playback experiment is convincing enough for me to accept the
split of the two forms. Plus, there are no other named taxa to consider.
K. NO. The “luteoviridis” complex includes
more than these two taxa and the relationships among them are likely more
complex than this study can establish. I suspect a proposal including the
phylogenetic work and voice analyses mentioned by Boesman would be sufficient
to consider, however
L. NO. Atlapetes no doubt have a number of surprises
left for taxonomists, but this case simply isn’t sufficient to sway me.
M. NO. Once again, there are far too many taxa within this complex for
this one study to suffice in judging how best to divide them up.”
Comments from Pacheco:
“A. YES.
Favorable from the available morphological, vocal and genetic data. Also, the "taxonomic inertia" mentioned
by Santiago.
B. YES. A
convincing split considering Schultz et al. 2017, with information obtained in
South America and more closely. The future may show a third taxon involved, as
Santiago alerts.
C. NO. It
lacks better genetic evidence and a better sampling of populations from
different subspecies located more closely.
E. YES.
Another case of "taxonomic inertia" remembered by Santiago.
Considering the genetic evidence (and others) mentioned by Boesman and
Santiago, I vote for this late split.
F. NO. The
separation in this case seems premature.
G. NO. The
data are suggestive, but the existence of gaps is evident.
H. NO.
Although suggestive, it is desirable that an analysis of the complex be carried
out with more geographic breadth.
I. NO.
Similar to the previous case. A better distributed sampling over the great
gradient of the distribution is desirable before a decision.
J. YES.
Vocal repertoire and distinct morphology with striking latitudinal separation.
I agree with Santiago: in this case, the burden of proof must fall on anyone
who wishes to demonstrate conspecificity.
K. NO. The
evidence for a split is still incomplete, mainly due to the gaps mentioned by
Dan.
L. YES. As
highlighted, the available genetic data demonstrate that these two taxa are not
even related.
M. NO. It
is premature to decide based on the information available.”
Comments from Robbins: “After reading Gary Stiles’s earlier and
more recent comments coupled with recent ones by Dan Lane, I’m now persuaded in
not accepting any of these proposals. Yes, for many of these, I suspect that
more than one species is involved, however, as Dan points out in several cases
other taxa need to be considered.”
Comments
from Stiles:
“The playback experiments of Freeman et al.: we can´t do anything here if
people keep voting NO “on principle”. My own feeling is that this is something
of a cop-out, for two reasons. First, I think that the birds themselves are the
best judges of whether a given recording is their species or something else. Therefore,
where these experiments were conducted with adequate replicates and clear
preferences were indicated (which I note was not always the case), they provide
important evidence regarding species status; this ESPECIALLY where other
evidence is available (including more information on vocalizations by Boesman,
plumage, genetics etc.) such that the playbacks might suffice to shift the
balance of proof to favor splitting the species in question, AND there is
precedent (at least, among the several most authoritative world lists) that the
split has already been made. Hence, I think that 3-5 of the 13-14 proposals are
sufficient to justify at least considering SACC action. So, over to the
committee.”
Comments
from Remsen:
“I acknowledge the validity of the points made against this approach by many of
you. If we were dealing with novel
splits in a species complex for which there were no previous classifications
that treated them as separate species or for which there was no indication in
the literature that species limits were questionable, then indeed I would not
endorse any of the proposals. However,
as pointed out by several of you, many of these splits were previously
anticipated or implemented based on anecdotal information. Others seek to rectify unsupported lumps from
the Lumperama Era. So, case-by-case
seems the best approach in my opinion.
Votes to follow “soon.”
Additional
comments from Stiles:
“I definitely agree with approval of B, J and L, and
could be persuaded on E (are there any genetic data for nigrita of
Venezuela??). I note that both playback and genetic data also support A).”