Proposal (838) to South American
Classification Committee
Treat Cinclodes
olrogi as conspecific with Cinclodes oustaleti
Effect on South
American CL:
This proposal would place Cinclodes olrogi within the Cinclodes
oustaleti species complex that includes various allopatric taxa:-
C.
o. oustaleti:
C Andes of Argentina and Chile south to N Patagonia
C.
o. hornensis:
S Patagonia and Tierra del Fuego
C.
o. baeckstroemii: Isla Alexander Selkirk formerly
Masafuera, Chile
C. [o.] olrogi:
Sierras of Córdoba and San Luis, CN Argentina
Background: Our current SACC note
reads as follows:
14a.
Nores (1986) considered Cinclodes olrogi to be a subspecies of C.
fuscus (broadly defined); others
(Olrog 1979, Navas & Bó 1987, Vuilleumier & Mayr 1987, Mazar Barnett
& Pearman 2001) considered it more likely to be closely related to C.
oustaleti. Genetic data support the latter relationship (Chesser 2004a,
Derryberry et al. 2011). See also Note
14b. SACC
proposal needed.
Cinclodes olrogi has long been known as
a controversial "endemic" species in Argentina ever since its
description in 1979. Different authors have classified it as either a distinct
species, as a subspecies of C. oustaleti or as a subspecies of C.
fuscus. The brief type description (Nores & Yzurieta 1979) treats olrogi
as a monotypic species although in the same year C. C. Olrog (1979) immediately
treated olrogi as a subspecies of oustaleti. It is also worth
noting that M. Nores subsequently changed his stance and considered olrogi
to be a subspecies of Cinclodes fuscus (Nores 1986, 1996). However, and
in the only exhaustive and critical morphological study of the oustaleti
complex, J.R. Navas and N. Bó (1987) convincingly demonstrated that olrogi
should be regarded as a subspecies of oustaleti based upon plumage and
morphometric data.
As invariably noted, virtually all of
the plumage features found in olrogi are present in either nominate oustaleti
or hornensis or both, and the white center of the abdomen is a shared
feature of all of these taxa (Fjeldså & Krabbe 1990, Navas & Bó 1987,
pers. obs.).
The concluding remarks by Navas and Bó
(1987) are also pertinent when they suggested that “the most important feature
[distinguishing olrogi from oustaleti] was a shorter mean
difference of 1-3 mm in the culmen of olrogi compared to oustaleti
and hornensis.”; Jorge R. Navas spent much of his life making meticulous
specimen comparisons as the MACN curator (Buenos Aires) of 60,000
specimens.
New information: A phylogenetic
analysis (Chesser 2004) based on mitochondrial DNA (complete COII and ND3
genes) showed a divergence of 0.5% between olrogi and oustaleti.
Chesser (2004) concluded that “it seems likely that they are potentially
interbreeding taxa, and they are perhaps best considered biologically
conspecific, pending further study.”
Using the same genes and partly the same
dataset, Sanín et al. (2009) recovered,
without much support, a sister relationship of olrogi and oustaleti,
but interestingly a haplotype of what is currently known as C. albiventris
tucumanus was more similar to oustaleti than to olrogi
(Figure 2). They also mentioned that “Although the status of the forms [of the
then C. fuscus] occurring in the central Andes and Argentinean highlands
is not entirely clear with respect to C. olrogi and C. oustaleti,
we suggest they should be maintained separate and elevated to the
species-level, taking the name C. albiventris. This species would
provisionally comprise five subspecies: C. a. albiventris, C. a.
tucumanus, C. a. yzurietae, C. a. riojanus,
and C. a. rufus. Additional work will be necessary to determine whether
these taxa are maintained as specifically distinct from C. olrogi and C.
oustaleti. (Sanín et al. 2009:554).”
Fig. 1. Chesser (2004): Phylogram of the ML tree
constructed using combined sequences of COII and ND3.
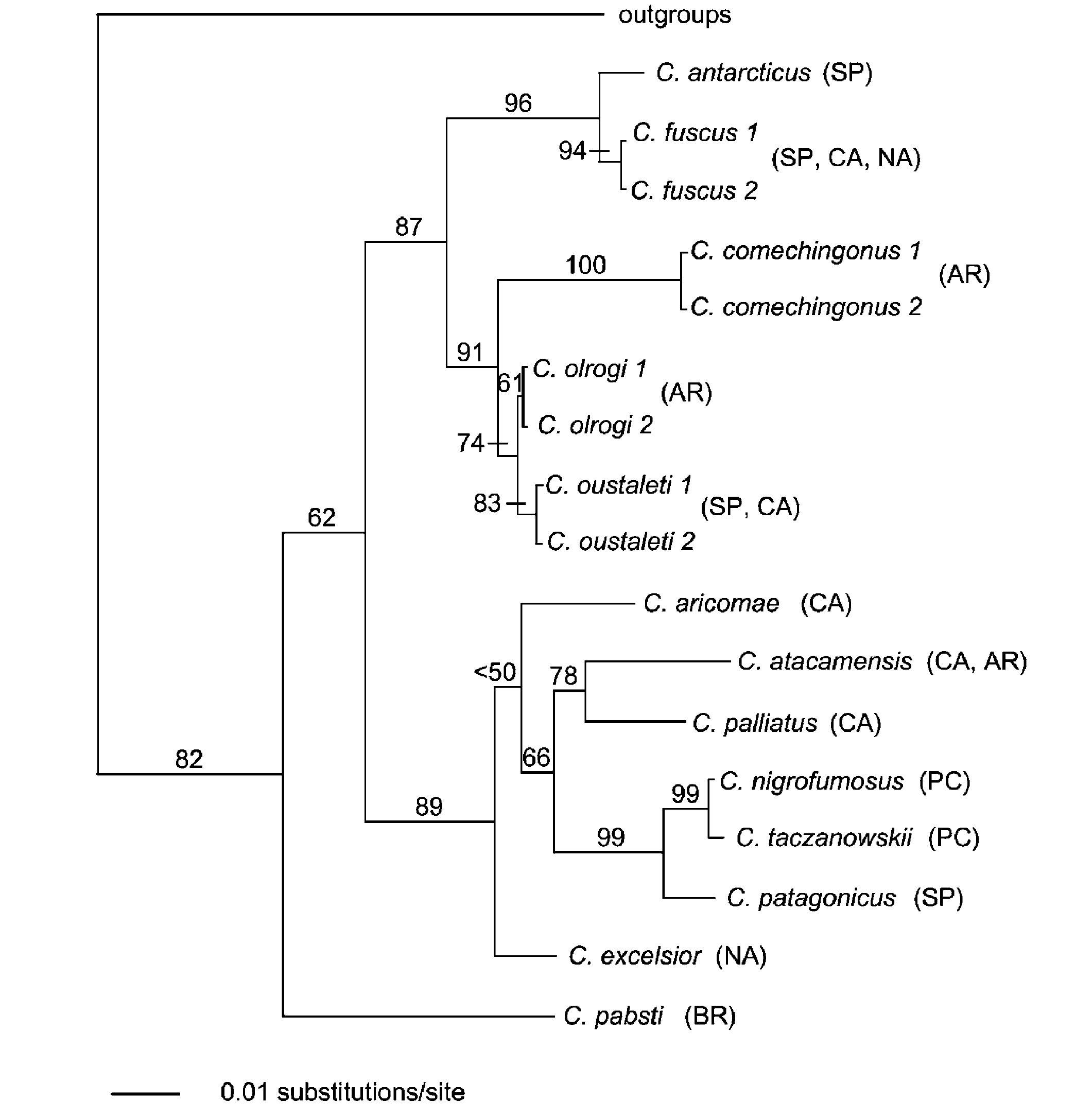
Fig. 2 . (Sanín
et al. 2009). Maximum clade credibility tree for ND3 and COII haplotypes
of C. fuscus and the other Cinclodes species, inferred using Bayesian analysis
as implemented in BEAST.
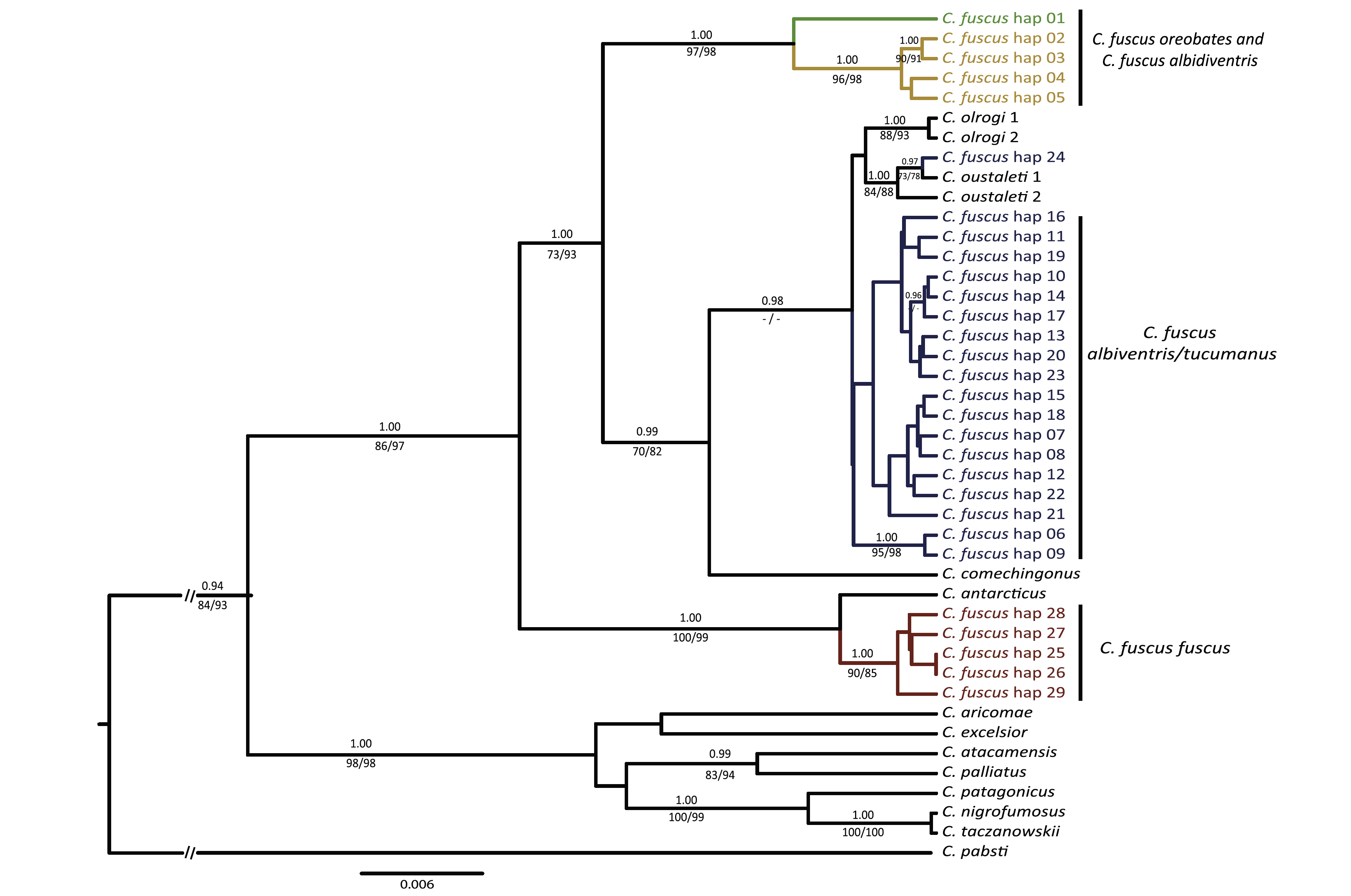
“Most haplotypes of the central
subspecies C. f. albiventris/ tucumanus formed a weakly supported clade
(0.59, 28%, 24%), which grouped with C. olrogi and C. oustaleti
(0.93, 40%, 42%). However, one haplotype, observed in two individuals from
Tucumán, Argentina, was identical to one of the two haplotypes of C.
oustaleti reported by Chesser (2004). In the Bayesian analysis, C. f.
albiventris/tucumanus was sister to a clade formed by C. olrogi and C.
oustaleti (posterior probability: 0.98), whereas in the MP and ML trees C.
oustaleti was weakly supported (<20%) as sister to a clade formed by C.
olrogi and C. f. albiventris/tucumanus. Divergence of some haploypes within C. fuscus albiventris/tucumanus
(range 0.09–1.06%) was greater than the divergence of haplotypes between C.
f. albiventris/tucumanus and C. oustaleti (range 0. 38–0.96%,
without considering the shared haplotype between them) or C. olrogi (range
0.29–0.87%). The haplotype network (Fig. 3) provided a different perspective on
relationships among these taxa. Although most C. f. albiventris/tucumanus
haplotypes clustered together, two haplotypes from central Peru were distant
from the rest. The two haplotypes of C. olrogi formed a cluster, as did
those of C. oustaleti (including the fuscus individuals that
share the haplotype), but these two clusters did not form a distinct unit.
These clusters were separated from the rest of the network by only two
mutations each, whereas the separate C. f. albiventris/tucumanus
haplotype cluster was four steps removed.” (Sanín et
al. 2009:551)
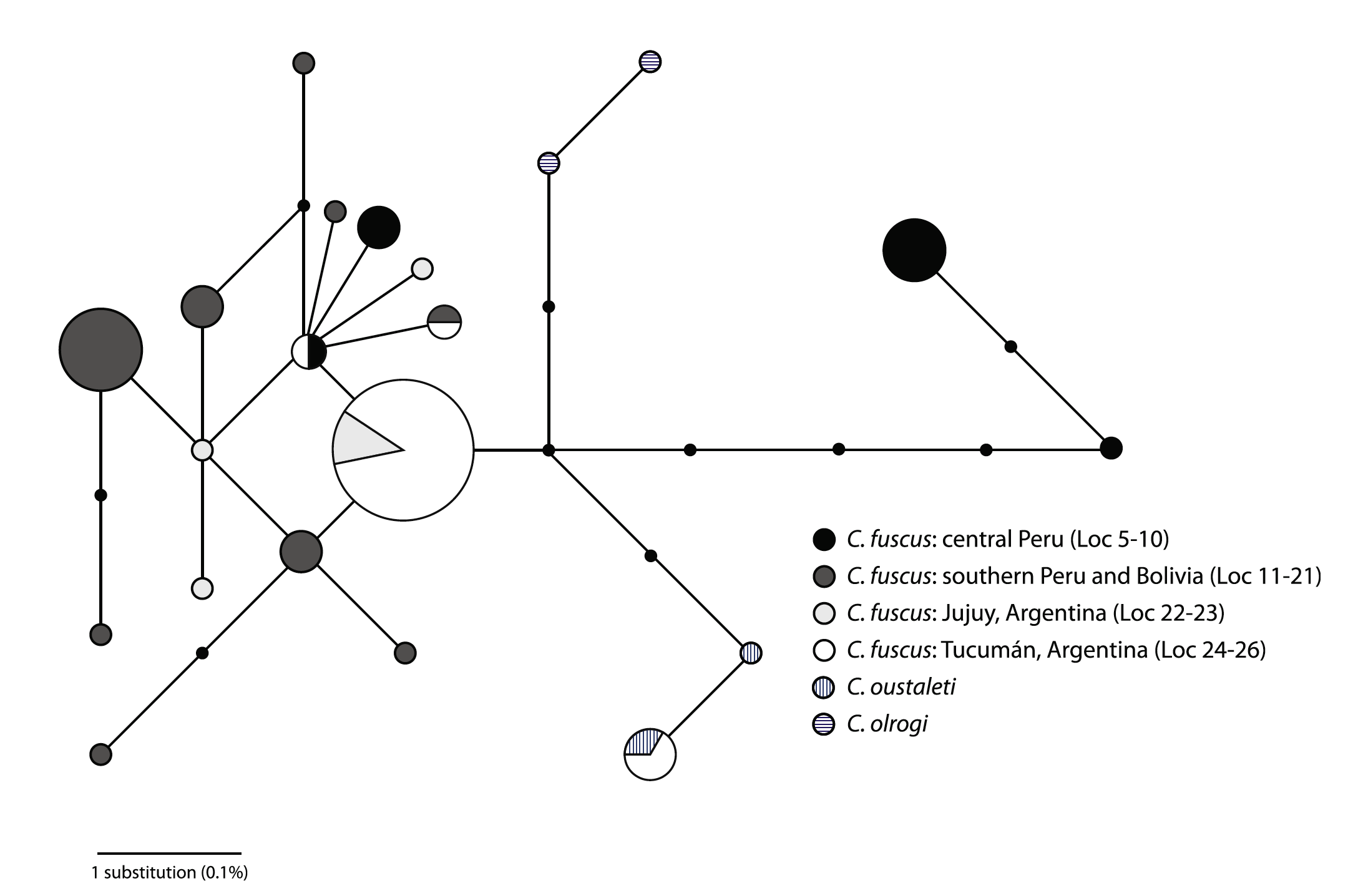
Fig. 3 (Sanín et
al. 2009). Median-joining haplotype network showing relationships among C.
fuscus albiventris, C. oustaleti, and C. olrogi haplotypes.
Sizes of the circles are proportional to the number of individuals sampled with
that haplotype. Connection lengths are proportional to the number of mutations
between two haplotypes. Shadings and patterns correspond to different regions
or species. Small black vertices represent unsampled or extinct haplotypes.
Analysis: Since Chesser (2004)
and Sanin et al. (2009), there has been a long period of silence, yet
published morphological and genetic data both support the notion that olrogi
is conspecific with oustaleti. There seem to be no lines of evidence at
all that would support recognition of olrogi as a valid species. This in
turn questions the recognition of albiventris as separate from oustaleti,
but more comparative material is needed to resolve this problem.
The taxonomic status of hornensis
and baeckstroemii in relation to oustaleti seemingly require further
study but are unrelated to this proposal.
All of these taxa are largely sedentary
with some north-south migration in hornensis, and also slight altitudinal
migration in oustaleti, hornensis and olrogi.
Recommendation: We recommend a YES
vote to lumping olrogi with Cinclodes oustaleti for the reasons
mentioned above.
Literature Cited:
Chesser, R.T. 2004. Systematics, evolution, and
biogeography of the South American ovenbird genus Cinclodes. The Auk
121 (3), 752-766.
Fjeldså, J. & Krabbe, N. 1990. Birds of
the High Andes: A Manual to the Birds of the Temperate Zone of the Andes
and Patagonia, South America. Zoological Museum, University of Copenhagen,
Navas, J.R. & Bó, 1987. Notas
sobre Furnariidae argentinos
(Aves, Passeriformes). Museo Arg. Ciencias.
Naturales 14(4): 55-86.
Nores, M. 1986. Diez nuevas
subespecies de aves provenientes de islas ecológicas argentinas. Hornero
12(4): 262-273.
Nores, M. 1996. Avifauna de la Provincia de
Córdoba. Pp 255-337 in Di Tada, I.E.
& Bucher, E. eds. Biodiversidad de la
Provincia de Córdoba, 1. Univ. Nac. Rio
Cuarto.
Nores, M. & Yzurieta, D. 1979. Una nueva especie y dos nuevas subespecies de aves (Passeriformes). Academia Nacional Ciencias Córdoba Misc. 61: 4-8.
Olrog, C.C. 1979. Nueva lista
de la avifauna argentina. Opera Lilloana 27.
Sanín, C., Cadena, C.D.,
Maley, J.M., Lijtmaer, D.A., Tubaro, P.L. & Chesser, R.T. 2009. Paraphyly
of Cinclodes fuscus (Aves: Passeriformes: Furnariidae): Implications for
taxonomy and biogeography. Molecular Phyl. and
Evol. 53: 547-555.
Mark
Pearman and Nacho Areta, July 2019
Comments from Claramunt: “NO. The arguments for
conspecificity presented are very weak and information on potential
reproductive compatibility is completely lacking:
“Plumage: Statements by
Navas & Bo, and Nores are based on “degree of similarity;” they concluded
that the differences are “at the subspecific level.” I admit that these species
are very similar (there is an obvious general plumage conservatism in Cinclodes
and other open country furnariids). Nothing strange there, especially given the
recent divergence times. However, overall similarity should not be interpreted
as evidence of conspecificity (or pheneticism is
back?). olrogi is fully diagnosable, at least by the shape and color of
the alar band, which is pure white and broad distally in olrogi
(conspicuous even in the closed wing), versus ochraceous and narrower in oustaleti.
This difference should not be downplayed, as it would be very conspicuous
during the singing display typical of this genus. Other plumage differences
such as the browner tones of olrogi may be also diagnostic.
“Morphometrics:
Differences in bill length are not tremendous but actually they show no
overlap: olrogi 14 to 16 mm (mean of 15 mm, 13 individuals), oustaleti
17 mm (9 individuals, no detectable variation). So, these species seem
diagnosable by bill length (at least no evidence of continuous variation).
“Genetics: The genetic
evidence only suggests that there may be some issue of incomplete lineage
sorting or past hybridization but between albiventris and oustaleti. C. olrogi is not involved in this
problem (it has its own, non-shared, haplotypes). So, no evidence of
conspecificity there.
“Genetic distances
between olrogi and oustaleti are similar to those between oustaleti
and albiventris, or between fuscus and antarcticus. So, no
evidence of anything there.
“Phylogeny: olrogi
appears as sister to oustaleti in the mtDNA tree, but the node that
unites them does not have statistical support, so we cannot rule out the
possibility that oustaleti is closer to albiventris, with olrogi
as a more distant relative.
“Therefore, I don’t see
any evidence of conspecificity between olrogi and oustaleti. Moreover, evaluating the status of olrogi
without considering the status of albiventris seems problematic.”
Comments from Remsen: “NO. I
agree with all of Santiago’s points above.
I would also add that I wonder if the phenotypic assessments of Navas,
Bó, and Nores would have been different if they had not been in the framework
of C. fuscus sensu lato (prior to Sanín et
al.).
“Even if this proposal
passed, it’s a good example of why any proposal that focuses the issues towards
what additional data are needed is productive (and more valuable than the
immediate outcome of the voting).
Specifically, what data are needed to assess species limits in this
genus? Are there any contact zones to
provide insight on barriers to gene flow that could be applied as a yardstick
to allotaxa (with all appropriate caveats)?
Are the vocalizations perceived by the birds as being as similar as they
do to us? Given that these are among the
most vagile furnariids, it seems likely that there are intrinsic barriers to
free gene flow.”
Additional comments from Pearman and
Areta:
“We would like to add some more information to
expand our proposal, because we feel that some of the arguments put forward by
Santiago demand clarification on our part.
“Plumage
“Santiago stated that "olrogi is fully diagnosable, at least by the shape
and color of the alar band, which is pure white and broad distally in olrogi (conspicuous even in the closed wing), versus ochraceous and narrower
in oustaleti. This difference should not be downplayed,
as it would be very conspicuous during the singing display typical of this
genus. Other plumage differences such as the browner tones of olrogi may be also diagnostic.”
“However, although we agree that olrogi
is diagnosable, his characterization may create a false impression of great
distinctiveness that is not borne in the field or in specimens. Diagnosability
does not equate to species status, and the critical point lies in understanding
what does diagnosability entails from the point of view of speciation. In our
proposal we did not enter into the finer detail of plumage distinctions because
these taxa are extremely similar and the proposal focused on the genetic data.
“Wing-stripes in Cinclodes can
be one color on the upperwing and a different color on the underwing, and upper
and underwing seem to differ on their importance during display. The
wing-raising display in all Cinclodes
involves a high, vertical wing spread with fluttering and rotation on an almost
vertical axis, meaning that the bird is always exposing the wing-flash or ulnar
bar of the underwing to a potential mate from an elevated perch; not the
upperwing.
“The underwing pattern of oustaleti
and olrogi is 100% identical in
coloration; a more recently collected olrogi
specimen was very slightly cleaner white than older oustaleti specimens. MP measured the maximum width of this ulnar
bar and found it to be 22 mm in both olrogi
and oustaleti which at least demonstrates
that from a single comparison they were identical, and not broader in olrogi as Santiago stated.

MACN specimens: C. oustaleti oustaleti (left) and C. olrogi (right)
“Moving on to the upperwing, the wing-stripe of both taxa is largely a
fairly bright cinnamon, with the difference that in olrogi the most distal portion becomes whitish; just barely visible
in the photograph below. This white portion can sometimes be seen on the closed
wing, but it is not used in display. Thus, both the upper and underwings of
these taxa are extremely similar. Importantly, Cinclodes oustaleti hornensis has an identical upper and underwing
pattern to olrogi.

MACN specimens: C. oustaleti oustaleti (left) and C. olrogi (right)
“We were surprised by Santiago´s suggestion that we are endorsing a
“pheneticist” point of view, because we are attempting to use whatever is known
of these taxa; and morphology seems to be the main source of characters at
stake. We submit that a small difference in bill length (not controlling for
the possible existence of latitudinal clines in oustaleti-hornensis) is meager evidence to grant species status to olrogi.
“Intraspecific clinal variation leading to mildly or wildly different
looking populations is known to occur along the Andes and neighbouring mountain
chains (e.g., Upucerthia dumetaria
and U. validirostris for which see
Areta & Pearman (2009, 2010), and the recent merger of Cranioleuca baroni and C.
antisiensis for which see Seeholzer & Brumfield (2017). We see that
available evidence for considering olrogi
as a different species from oustaleti
is exiguous, whereas considering them as conspecific is more consistent with
available data on morphology and genetics. On a similar vein, Cinclodes atacamensis schocolatinus
(from the Sierras de Córdoba and San Luis) is as different from the Andean Cinclodes atacamensis as olrogi is from oustaleti. Yet, nobody seems to be arguing for a split here.
Interestingly, in both cases the Córdoba populations are darker than the Andean
populations.
Genetics
“We do not advocate for a genetic-threshold for species recognition, and
are aware that good biological species can differ minimally in this regard.
However, we fail to see how having two slightly different haplotypes provides
solid evidence of the existence of two species per se. What we argued is that the genetic data is weak, and thus
it should be interpreted with care. The fact that “genetic distances between olrogi and oustaleti are similar to those between oustaleti and albiventris”
is put forward as an argument, but it can instead be used to undermine the
recognition of albiventris as
distinct from olrogi and oustaleti. The issue of whether albiventris is conspecific or not with olrogi-oustaleti is a lingering one.
Interestingly, the taxa yzurietae-riojanus-famatinae are but
reddish versions of oustaleti-olrogi,
while albiventris is a browner
version sharing the same structural plan.
“The lack of support for the “albiventris”
group (see Figure 2) and the sharing of haplotypes of albiventris with oustaleti
suggest that the story is not simple, yet, the split of albiventris was accepted without serious conflict on this topic by
SACC (http://www.museum.lsu.edu/~Remsen/SACCprop415.htm). Recognizing the extremely similar and genetically weakly
differentiated olrogi and oustaleti as a single species can be
considered as a necessary first step to solve the taxonomy of this group.
“At any rate, genetic evidence can be interpreted in many difference
ways. From our perspective, however, the most difficult interpretation is that
the scant genetic differentiation provides evidence in favou\r
of the recognition of two species. It can be consistent with two species, given
that some species exhibit almost null genetic differences, but it is difficult
to propose it as a solid argument in favor of species status for olrogi.
Vocalizations
“Vocal differences, if any, remain to be shown. In the few recordings
available, vocalizations of all taxa in the albiventris
complex are very similar. More recordings are needed, because individual birds
can change the pattern of song within the same bout (pers. obs.). So caution is
advised when jumping quickly to conclusions regarding differences based on a
few recordings.
Final remarks
“Although Santiago routinely mentioned “no evidence” indicating
conspecificity of olrogi and oustaleti, we argue that evidence
supporting their recognition as different species does not fit well with
current species limits in Cinclodes.
To this end, it might be relevant to bring back the case of Cinclodes espinhacensis (which was
considered to be subspecies of C. pabsti
by this committee: http://www.museum.lsu.edu/~Remsen/SACCprop548.htm), and the problematic C.
nigrofumosus/taczanowskii duo, which seem to be more a sign of historical
momentum than of solidly sustained biological species (http://www.museum.lsu.edu/~Remsen/SACCprop33.htm).
“In sum, this is not an easy case, but C. olrogi is a little-differentiated taxon both in morphology and plumage, vocalizations
resemble those of C.
oustaleti/albiventris and it shares a distributional pattern with
subspecies schocolatinus
of C. atacamensis in the Sierras de Córdoba. This, added to the nature of
geographic variation in Cinclodes
(and other Andean furnariids, including the closely related Upucerthia) should put the burden of
proof in those advocating full species status for olrogi.”
Additional comments from Remsen: “I am swayed by the new comments
from Mark and Nacho, and I change my vote to YES. More broadly, the issue of species limits in
this genus really needs re-evaluation, especially from the standpoint of voice
and playback trials.”
Comments from Zimmer: “YES”. I
must confess to being influenced in this case by my field experience with Cinclodes espinhacensis and C. pabsti (which seem to differ at least as much in
plumage characters as olrogi and oustaleti), which showed 2 pairs of espinhacensis to respond instantly,
vigorously and repeatedly to playback of both single-note calls and songs from
multiple pabsti (see my detailed
comments to Proposal #548). This
experience not only caused me to change my vote on Proposal #548, but it also
left me feeling less than confident about the morphological “yardsticks” used
for evaluating species-limits within this evolutionarily conservative genus.”
Additional comments from Claramunt: “This case is not similar to the
espinhacensis case and the discussion of the wing patch by Nacho and
Mark is misleading: 1) the dorsal side of the wing is clearly visible during
displays in Cinclodes, 2) specimens in the pictures are inadequate to show
anything useful as the difference between oustaleti and olrogi is
in the distal section of the wing, in the primary feathers that are concealed
in the specimens (not in the “ulnar” = secondary feathers). Regarding
morphometrics, I have measured specimens of both species and, although sample
sizes are mall (n=3, 4) I can confirm the bill size differences using bill
length from nares (olrogi 10.6-11.4 vs. oustaleti 12.2- 13.3mm)
and I realized that they also differ in hallux length with no overlap (olrogi
16.2-17.2 vs. oustaleti 18.5- 19.4mm). So, this is different from the
case of pabsti/espinhacensis, which show wide overlap in all
measurements. Instead, olrogi shows diagnostic differences in at least
one plumage trait and two morphometric traits. Only a phenetic point of view
would disregard these differences on the ground of being of small magnitude.
Instead, I think that these differences demonstrate a history of reproductive
isolation that explains the accumulation of diagnostic differences between
these two taxa.”
Comments
from Bonaccorso:
“NO. Driving away from
the “pheneticist” discussion, lack of genetic divergence is just one line of
evidence. Both plumage and morphological differences seem subtle, but are
discrete. Also, it is important to evaluate plumage differences in the context
of subtle plumage differences found among other species of Cinclodes
(e.g. C. fuscus and C. albidiventris). I have not seen C.
olrogi displaying, but if the white wing patch is conspicuous during
display, it is definitively a key feature that could point to reproductive
isolation. Also, even if future analyses confirm that Mark and Nacho are right,
it will be easier to just lump these two taxa. However, if future analyses show
that they are not conspecific, we will have to reverse the lumping, creating
more chaos.”
Comments
from Robbins: “YES. The minor level of differentiation in both
morphological and genetic data of olrogi suggest that it should be
treated as conspecific with oustaleti.
Yes, there needs to be a re-evaluation of species limits within this entire
genus. I vote for subsuming olrogi
into oustaleti.”
Comments from Stiles: “In view of the most interesting
(and confusing, at least to me) discussions on the olrogi-oustaleti
question, I think it best to abstain (at least for now). It seems that the same
genetic and plumage data can produce divergently opposite opinions regarding
the question of the proposal itself ... and looking at the Sanin genetic data,
it appears that the olrogi-oustaleti question is embedded within
the wider question of species limits in albiventris vs. fuscus.
It looks to me like a question of back to the drawing board - i.e., the birds
in the field. Given that plumage differences appear subtle, at least to my
uneducated eye, vocalizations and displays, coupled with extensive reciprocal
playback experiments, may be the way to go (would make a nice doctoral
thesis?).”
Comments from Pacheco: “In view of the difficulty in
deciding from the information available at this moment, I choose to abstain.”
Comments from Jaramillo: “NO. I am most swayed by Santiago’s viewpoints on
this problem. But also I will note something pretty remarkable about C.
oustaleti, at least the nominate population. They are migratory; now, the
situation might be a tad different on the Argentine side, but in Chile the
majority move to the coast in winter. What is striking is that they do not just
move “coastward” but they setup immediately in the splash zone of the coast and
share the habitat with C. nigrofumosus (Seaside Cinclodes). This strikes
me as a particularly amazing shift in habitat for the majority of the
population, going from a High Andean breeding habitat to a marine habitat. This
is possible through physiological adaptations that are similar to that of C.
nigrofumosus. Apart from a move to marine habitats, there is a slight
northward movement of oustaleti compared to the breeding range. Here is
one of the papers on the physiology of oustaleti.
https://link.springer.com/article/10.1007/s00360-004-0428-2
“I think that apart from the missing
details mentioned by Santiago in the discussion. It is important to note this
quite different ecology, at least of the major proportion of the nominate
subspecies of oustaleti. This situation of some taxa of Cinclodes
being marine or partly marine is interesting, and it has occurred in different
branches of the Cinclodes tree. Additionally, something unusual may be
happening in the far south, with oustaleti. It appears that there are
Andean breeders and perhaps marine breeders, within oustaleti. Which one
of these, or both, refers to hornensis might have bearing on oustaleti
(sensu stricto). But indeed this, and the Juan Fernandez population do not have
direct bearing on the question at hand.
“I
am open to the idea that these taxa may be conspecific, but I think we are not
quite there in determining that.”
Comments from Stotz: “NO.
I am uncertain what the best treatment here is; given my uncertainty, I am
inclined to continue with the status quo.”
Comments
from Pacheco:
“NO. I took a closer look
at the arguments on both sides: Pearman / Areta versus Santiago and chose to
consider the same cautious position as Elisa. Despite genetic similarity, the existence of
an objective diagnosis (without overlapping) and evidence of a potential
reproductive isolation make me decide to keep both taxa at the species level.”