Proposal (841) to South American
Classification Committee
Change the species
status of two swifts of the genus Chaetura:
(A) recognize C. andrei as a valid
species, and (B) treat C. viridipennis as
conspecific with C. chapmani
Background:
Chaetura andrei was described by
Berlepsch and Hartert in 1902 based on specimens from Venezuela. Hellmayr later described C. andrei meridionalis as a subspecies of andrei and most subsequent authors (e.g., Cory 1918, Peters 1940,
Meyer de Schauensee 1966) considered the two forms to be conspecific. Marín
(1997), however, argued that andrei is
indistinguishable from C. vauxi aphanes and
recommended that the two forms be considered the same taxon (the name andrei has priority), as a subspecies of
vauxi. He also recommended that meridionalis be recognized as a separate
species.
Chaetura viridipennis was described by
Cherrie in 1916, based on a single specimen from Mato Grosso, Brazil, as C. chapmani viridipennis. Cherrie noted that it is similar to chapmani but “larger and differs in
having the pileum, mantle and wings glossed with greenish (instead of
steel-blue).” Cherrie’s treatment of viridipennis as a subspecies of chapmani was followed by most if not all
authors (e.g., Cory 1918, Peters 1940, Meyer de Schauensee 1966), until Marín
(1997) recommended that chapmani and viridipennis be considered separate
species based on differences in wing length similar to the differences between C. vauxi and C. pelagica.
SACC
adopted the recommendations of Marín (1997), treating andrei as a form of vauxi,
treating meridionalis as a species
distinct from andrei, and treating viridipennis as a species distinct from chapmani.
New Information:
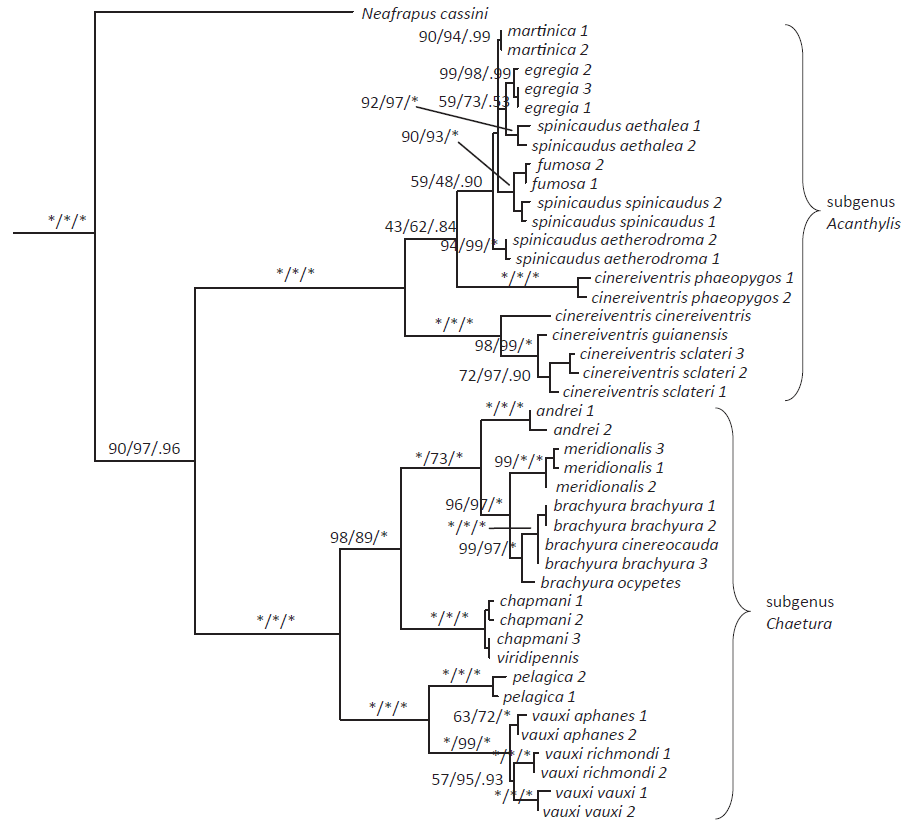
Chesser
et al. (2018) sequenced mitochondrial and nuclear DNA for all species and many
subspecies of the genus Chaetura,
including andrei, meridionalis, vauxi aphanes, chapmani,
and viridipennis. Their results (see
tree below) indicated that andrei is
not closely related to vauxi aphanes,
and that it is quite distinct from meridionalis
as well, being sister to the clade consisting of brachyura + meridionalis.
Mean mitochondrial divergence between current conspecifics andrei and vauxi aphanes
was 7.0%, and divergence between former conspecifics andrei and meridionalis
was 3.2%, comfortably within the range of divergence between species in this
genus. Chesser et al. (2018) also compared skins of andrei and vauxi aphanes,
and concluded the following:
“Plumages of the
two taxa are quite distinct: the lower breast and belly of andrei are noticeably darker than those of vauxi aphanes, and the undertail coverts of andrei are paler than or concolorous with the belly, whereas the
undertail coverts of vauxi aphanes are darker than the belly.
Moreover, the light area of the throat tends to be smaller and better
delineated in andrei than in vauxi aphanes, and the upperparts of andrei
are lighter brown (olive brown) than those of vauxi aphanes, which are blackish-brown. Even worn plumaged vauxi aphanes were readily
distinguishable from andrei.”
In
contrast, Chesser et al. (2018) found chapmani
and viridipennis to be virtually
identical genetically, closer even than some individuals within subspecies.
Mean mitochondrial divergence between the two forms was 0.1%. This supports the
conclusion of Collins (1968), who found that the color differences in
iridescence between these two taxa are related to degree of feather wear and
not indicative of taxonomic distinctiveness. Thus, the only valid published
morphological difference between chapmani
and viridipennis is in wing
length, as would be expected for sedentary (chapmani)
and migratory (viridipennis) forms of
the same species.
Recommendation:
As in
Chesser et al. (2018), I recommend (a) that andrei
be recognized as a species distinct from current conspecific C. vauxi aphanes and former conspecific C.
meridionalis, based on its phylogenetic and phenotypic distinctiveness from
these and all other forms of Chaetura.
This would also reinstate aphanes as
a recognized subspecies of C. vauxi.
In addition, I recommend (b) that viridipennis
be returned to its former status as a subspecies of C. chapmani, based on the combined genetic and phenotypic data.
Although it may be subspecifically distinct from chapmani, viridipennis
differs only slightly from that form in both genotype and phenotype, and does
so to a much lesser degree than other Chaetura
taxa generally considered specifically distinct.
Lit Cited:
Cherrie, G. K. 1916.
Some apparently undescribed birds form the collection of the Roosevelt South
American expedition. Bull. AMNH 35: 183-190.
Chesser, R. T., H. Vaseghi, P. A. Hosner, L. M. Bergner, N. Cortes-Rodriguez,
A. J. Welch, and C. T. Collins. 2018. Molecular systematics of swifts of the
genus Chaetura. Molecular Phylogenetics and Evolution 128: 162-171.
Collins, C. T. 1968. Notes on the biology of Chapman’s Swift Chaetura
chapmani (Aves, Apodidae). Amer. Mus. Novitates 2320: 1-15.
Cory, C. B. 1918. Catalogue of birds of the Americas. Field Museum of
Natural History Publications, Zoological Series, vol. 13, Part 2, vol. 1.
Marín, M., 1997. Species limits and distribution of some New World
spine-tailed swifts (Chaetura spp.). Ornithological Monogr. 48: 431–443.
Meyer de Schauensee, R. 1966. The species of birds of South America and
their distribution. Academy of Natural Sciences, Philadelphia.
Peters, J. L., 1940. Check-list of birds of the world, vol. IV. Harvard
Univ. Press, Cambridge, Mass.
Terry Chesser, November 2019
Comments
from Areta:
“I expect extant type specimens to be
critically examined in taxonomic revisions, especially in a group such as
swifts in which differences are minor and have led to a myriad of treatments
and proposed relationships. I also expect to see pictures of specimens when
differences are not obvious or clear cut, and to find relevant information such
as date and coordinates of specimens reviewed in the paper itself. Neither
Chesser et al. (2018) nor Marin (1997) fulfilled my expectations in these
regards.
A. Perhaps Yes. But I would like
more information. It makes me uneasy to recognize this species without any
information relating to its life (no breeding data, no known recordings of the
vocalizations of andrei), but I
cannot imagine any other reasonable option to sort out the taxa than this.
Despite this, I would like to see a
comparison between the type specimen of aphanes
(from Caripe) against the type specimen of andrei
(from Caicara). I must also point out the existence of a specimen identified as
andrei in the original description by
Berlepsch & Hartert (1902) and by Marin (1997) from San Félix in the
Turimiquire massif (referred to as Eastern Coastal Range by Marin), which if
correct, would suggest the possibility that aphanes
and andrei are the same thing (indeed,
this non-Orinocan specimen is the one responsible for the taxon
being named after E. André, who just sent the specimen to the Tring Museum,
although it was not designated as type specimen). Without careful examination
of type specimens of both taxa, we cannot be sure of what is going on there.
The samples of presumed aphanes used by Chesser et al. (2018) could have
pertained to richmondi instead, and
Marin (1997) might have been right in suggesting that andrei and aphanes are
but the same bird.
“Also, there is some plumage variation that has not been
adequately linked to genetic samples or to locality:
“1) Marin (1997) stated: ‘Two of the
known specimens of C. a. andrei, including the holotype
(AMNH 477325), show strong contrast between the throat and chest, whereas
the other two show less contrast. Phenotypically, the two individuals of C.
a. andrei that have less contrast are identical to the
southernmost form of C. vauxi [richmondi] aphanes.’ Which specimens
are those?
“2) Chesser
et al. 2018 wrote: ‘Plumages of the two taxa are quite
distinct: the lower breast and belly of andrei
are noticeably darker than those of vauxi
aphanes, and the undertail coverts of andrei
are paler than or concolorous with the belly, whereas the undertail coverts of vauxi aphanes are darker than the belly. Moreover, the light area of the
throat tends to be smaller and better delineated in andrei than in vauxi aphanes, and the upperparts of andrei are lighter brown (olive brown)
than those of vauxi aphanes, which
are blackish-brown. Even worn plumaged vauxi
aphanes were readily distinguishable from andrei.’ How did the samples
that were sampled for genetics look? Did they look like the type as assessed by
Marin (1997), did they differ from it, or was there one with contrasting throat
and one with less contrasting throat? Where do the aphanes specimens (AMNH 150208–150209, AMNH 150211,
AMNH 648819, and AMNH 786081–786083) used for comparison against andrei came from? From the type locality? From the Turimiquire
massif/Eastern Coastal Range (perhaps true aphanes?)
or from the Coastal Cordillera/Western Coastal Range (perhaps richmondi?)
“B. Perhaps Yes. Again, I see no
mention of comparison of the type specimen of viridipennis (which was described based on a single specimen)
against specimens sampled for genetic analyses. How can we be sure that the
samples attributed to viridipennis
are indeed representative of the type specimen without a comparison? Just based
on locality? With the high mobility of swifts, I would not use locality as
indicative of much. Especially in a taxonomic revision and given the migratory
behavior of this taxon.”
Comments
from Robbins:
“NO. For the very good reasons outlined by Areta, I vote no until these issues
can be addressed.”
Comments from Stiles: “NO, for the moment. Although I think that
Chesser is probably right on both points, the devil is in the details: the
relations between plumages, distributions and genetics need to be better
defined, including at least photos of the type specimens involved (if these
were not genotyped). In fact, a good series of color photos in both the vauxi
aphanes-andrei group and the meridionalis-chapmani
group (the latter showing the range of color glosses in each taxon) would be
most helpful, including giving which specimens were genotyped and which not and
their collection localities.”
Comments from Bonaccorso: “YES to A and B. The phylogeny supports Chaetura andrei as
a valid, evolutionary independent entity, and there are morphological
differences that allow its diagnosis and separation from species in its sister
clade. On the contrary, C. chapmani and C. viridipennis should be
conspecific because of lack of both morphological and genetic differentiation.”
Comments from Chesser: Response to Areta (1-5) and Bonaccorso (6-7)
comments on Proposal 841:
“1. Chesser et al. (2018) is not a taxonomic revision
– it is a molecular systematics study, as indicated by the title “Molecular
systematics of swifts of the genus Chaetura”. Some of our molecular results were at odds
with SACC taxonomy, and two of these were supported by plumage data. We would all like to have more information on
the behavior of these species, but for birds like swifts for which behavioral
data can be scarce, the combination of genetic and morphological data can be
persuasive, which is why we recommended changes to the taxonomy.
“2. The samples of aphanes in Chesser et al.
(2018) were from Venezuela and could not have been richmondi, the
distribution of which is Mexico south to Panama (BNA, HBW, etc.). At one time, the range of richmondi was
considered to extend south to Venezuela, but this was before Wetmore and Phelps
described aphanes in 1956. We did
sample richmondi (from Costa Rica) and these samples clustered together,
separate from our samples of aphanes from Venezuela. All individuals of richmondi and aphanes
formed a clade with our samples of nominate vauxi, exactly as would
have been expected.
“3. At the AMNH, I directly compared 4 specimens of andrei
(many or most of the known skins, including the type) with 7 specimens of aphanes,
all of which were from localities either mentioned in the description of aphanes
or very close by: El Limon, Valley of Puerto la Cruz (150208); Las Trincheras,
Estado Carabobo (150209); Boqueron, Estado Falcon (150211); Miranda, Carupao
(648819); and Rancho Grande, Portachuelo Pass (786081-786083). Differences between these taxa were obvious
and consistent, as described in Chesser et al. (2018) and below. Furthermore, the characters that distinguish
the AMNH series of aphanes are also apparent in the type specimen (USNM
461693), which I have examined. These
are not the same taxon.
“4. Both Charlie Collins (in 1998) and I (in 2016)
examined specimens of andrei at the AMNH in light of Marín’s (1997)
statement that aphanes and andrei are the same taxon. The 4 specimens of andrei examined
were 477325 (the type, from Caicara), 477326-477327 (from Altagracia), and
477328 (from San Felix). The characters that most readily distinguish andrei
from aphanes (much darker lower breast and belly of andrei
compared to aphanes, undertail coverts lighter than belly in andrei
vs darker than belly in aphanes, rectrices of andrei narrower than those of aphanes, and
middle back and crown medium to olive brown in andrei vs blackish-brown in aphanes) differ consistently between specimens of the two
taxa. Moreover, Miguel Lentino (pers.
comm.) points out (and he has examined specimens both at the AMNH and in
Venezuelan collections) that andrei has a whitish superciliary that is
lacking in aphanes. This difference,
among others, is shown in Restall's illustration in vol. 2 of Birds of
Northern South America (p. 223), where the authors state the following
regarding the lump of andrei: “We do not accept this, as it is apparent
from comparative examination in COP that they are distinct, and they have
different habits and habitat.” Charlie,
Miguel, and I have all determined that andrei consistently differs in phenotype from aphanes, differences also very apparent in the genetics. Regarding Marín’s remarks on ventral plumage,
there is some variation in contrast between throat and breast in the
series of andrei at the AMNH,
ranging from somewhat delineated in 477326 and 477327 to well delineated in
477325 and 477328. The individuals that Marín considered identical to aphanes
are presumably 477326 and 477327, both of which were sampled for the
genetic study; however, although these individuals are closer to aphanes
in this character than are 477325 and 477328, they are not identical, nor do
they resemble aphanes in the other characters noted above that more
readily distinguish the two taxa.
“5.
Our genetic sample of viridipennis was collected 11 Nov. 1986 at
Cachoeira Nazaré, Rondônia, between two of the few
previous localities of this form (in Mato Grosso and Acre). It seems
reasonable to conclude that this bird was viridipennis, given that it was collected within the range of
viridipennis and well out of the range of
nominate chapmani (and given that chapmani,
unlike viridipennis, is not known to be migratory). Moreover,
results of the Cachoeira Nazaré expedition were published (Stotz et al. 1997), and
an individual ca. 1000 km or more out of its known range but within the range
of a different subspecies might have been expected to be noted in the
publication, which it was not.
Only the tissue from this trip was exported to FMNH; the specimens went
to the MPEG. Their label identifies the
skin (MPEG 40222) as viridipennis and the wing measurement of this bird
(125 mm) is close to that of Cherrie's type (127 mm). These measurements fit in well with those of
specimens of viridipennis here at USNM (125-130; n=3) and are greater
than any of our nominate chapmani (115-122, n=6) and the 120 provided
for chapmani by Hellmayr (the flattened wing measurements of Marin are
somewhat longer for all). We had better
geographic coverage of chapmani/viridipennis than for most species,
including one sample from extreme western Guyana, one from eastern Guyana, one
from north-central Amazonia, and one from southwestern Amazonia. The lack of
genetic distinctiveness among these samples is consistent with the lack of
distinctiveness between chapmani and viridipennis in plumage
(Collins 1968).
“6. The taxonomic changes in SACC 841 were proposed precisely
because the genetic and plumage data are in agreement. Other possible changes based on Chesser et
al. (2018), such as a split of the genetically extremely distinctive cinereiventris
phaeopygos, for which we know of no non-molecular support, have not been
proposed because of the lack of phenotypic data. If I were considering this proposal, then I
would also bear in mind the strength of the case supporting the original lump
of andrei and split of viridipennis, both of which were novel
(and in the case of andrei, radical) taxonomic changes based on faulty
(as with andrei) or slight (as with viridipennis) morphological
data, with no corroborating information.
“7.
Support for our tree is excellent for most nodes and quite strong overall, and
support for the node uniting andrei and for the node uniting chapmani
and viridipennis is 100% bootstrap/1.0 posterior probability across
the board, as indicated by the asterisks above the branches in the phylogeny.”
Additional comments from
Areta: "I thank Terry
for his detailed comments. Regardless of the title, the paper touches deeply on
the taxonomy of Chaetura swifts, and his reply confirms that this
evidence, necessary to make taxonomic decisions, was missing from the paper.
With the details on the geographic provenance of specimens, how they looked,
and the examination of type specimens being put in perspective, I change my
votes from "Possibly Yes" to "YES"."
Additional comments from
Stiles: "Given the full
discussion by Terry of the genetic and phenotypic bases for his split of aphanes and
lumping of chapmani and viridipennis, I agree with Nacho that YES
is fully justified for this proposal.”
Comments from Pacheco: “A and B. YES. After Terry's
clarifications, I agree that the two recommendations are well supported.”
Comments
from Claramunt: “A. YES. B.
NO. If viridipennis is diagnostic by wing length and migratory (versus
the sedentary chapmani). I would like to see more evidence before
lumping based on little genetic differentiation alone.”
Comments from Jaramillo: “A.
YES. It appears to me that there is a species-level taxon here. I am convinced
from Terry Chesser’s notes that this is andrei,
and that specimens of that taxon (including type) differ from aphanes. I
think that Nacho’s comments were incredibly helpful, but Terry’s reply set my
mind at ease on several of these points.
“B. YES.
It seems that burden of proof is on keeping them separate. I am not even clear
that viridipennis is migratory from what I have seen, it is suggested
but I do not know . No data exists as to where viridipennis breeds, as
such we really don’t even know what its distribution actually is. Maybe I am
wrong, but it does not seem like there are many specimens of this taxon. I am
not quite clear that the wing length differences are something we can have much
trust in, given the low sample sizes. Are they clinal, sex linked, age? Who
knows? As such and given that we really have nothing to hang a hat on for this
form, and the molecular data show that it is marginally different from chapmani,
I think the only option is to lump it.”
Comments
from Zimmer:
“YES” to both A and B. The recognition of andrei as a species seems well supported by two data sets, and I’m
not sure there are any hard data to support the continued recognition of viridipennis as being distinct from chapmani.”
Comments solicited from Dan Lane: “B. This split really never made much sense to me (and the resulting
English names given to C. viridipennis have always made me cringe:
"Amazonian" or "Matto Grosso." Yech). The Chaetura are a pretty conservative bunch,
but a few characters seem to be important: rump, vent, throat, and chest
shades, and voice. In the case of most Chaetura, voice is actually a
great way to separate them, even allospecies such as C. egregia and C.
spinicauda have distinctive voices from one another. I cannot find any
useful distinctions between C. chapmani and C. viridipennis. In
fact, other than wing length, there seem to be no really good characters to
separate them. I am a bit skeptical of the claim of "migrant" status
for viridipennis (how was this determined, and by whom?), but the two
taxa appear to be allopatric (with the caveat that this may be largely circular
reasoning reinforcing that supposed status) and otherwise are nearly
indistinguishable genetically and by plumage. As a result, Terry's conclusions
sit well with my own world view of chapmani and viridipennis.
Lump 'em, I say! YES.”
Comments from Stotz:
“A. YES. Given the
distance on the tree between C. andrei and any of the vauxi, plus
the fact that C. vauxi is a tight group otherwise makes this split seem
straightforward,
“B. YES. Never fully
understood the split of viridipennis from chapmani. Given the limited morphological and genetic
distinctions between these two taxa I support returning to the Meyer de Schauensee
taxonomy.”
Comments
from Robbins:
“After reading Chesser's newly posted comments, I'm now persuaded to vote
yes for both A (recognizing andrei as a species) and b (treating viridipennis
as conspecific with chapmani).”