Proposal (856) to South American Classification Committee
Resurrect Cyanophonia
and revise the linear sequence of Euphoniinae (Fringillidae)
Effect
on SACC:
A “YES” vote on this proposal will resurrect Cyanophonia and place Euphonia
cyanocephala, E. elegantissima, and E. musica within it. The
linear sequence of species within Cyanophonia and Euphonia would
also be revised to reflect new information on phylogenetic relationships.
Background
and New Information:
Prior
to the recent publishing of a phylogenomic study of the true-finch subfamily
Euphoniinae, the most comprehensive molecular phylogeny of the group to date
(Zuccon et al. 2012) found that Euphonia was paraphyletic with respect
to Chlorophonia, the other genus in the subfamily. Specifically, Euphonia
musica was found to be sister to Chlorophonia cyanea. The authors
noted this taxonomic conflict but suggested that further sampling of species
would be necessary to adequately resolve this issue. More recently, Imfeld et
al. (2020) used target-capture and enrichment to sequence thousands of
ultraconserved elements and mitogenomes for every Euphonia and Chlorophonia
species presently recognized by the checklist committees of the American
Ornithological Society. Every phylogenetic analysis performed in this study,
whether with concatenation or species-tree methods, maximum likelihood or
Bayesian approaches, or using nuclear or mitochondrial sequence data, found the
three species of blue-hooded euphonias (Euphonia cyanocephala, elegantissima,
and musica) to form a monophyletic clade sister to the 5 Chlorophonia
species to the exclusion of all other euphonias (see figures and tables
below).

Table
1. Support metrics for the time-scaled phylogeny. This table is a pruned
version of Table 2 from Imfeld et al. (2020) and corresponds to the node
numbers in Figure 1 above. Abbreviations: localized posterior probability
(LPP), gene concordance factor (gCF), site concordance factor (sCF), and
internode certainty (IC).
|
Node |
LPP |
gCF |
sCF |
IC |
Bootstrap |
|
1 |
1 |
77.2 |
89.0 |
0.614 |
100 |
|
2 |
1 |
19.5 |
47.2 |
0.136 |
100 |
|
3 |
1 |
18.1 |
50.6 |
0.204 |
100 |
|
4 |
1 |
30.8 |
70.2 |
0.377 |
100 |
|
5 |
1 |
10.9 |
35.0 |
0.000 |
100 |
|
6 |
1 |
27.2 |
58.9 |
0.278 |
100 |
|
7 |
1 |
38.8 |
70.8 |
0.353 |
100 |
|
8 |
1 |
29.5 |
41.7 |
0.025 |
100 |
|
9 |
1 |
36.8 |
70.5 |
0.392 |
100 |
|
10 |
1 |
57.6 |
73.9 |
0.358 |
100 |
|
11 |
1 |
48.8 |
79.5 |
0.634 |
100 |
|
12 |
1 |
42.5 |
59.0 |
0.114 |
100 |
|
13 |
1 |
18.5 |
47.7 |
0.093 |
100 |
|
14 |
1 |
23.7 |
50.0 |
0.116 |
100 |
|
15 |
1 |
52.6 |
86.4 |
0.510 |
100 |
|
16 |
1 |
27.7 |
47.7 |
0.054 |
100 |
|
17 |
1 |
28.8 |
50.1 |
0.041 |
100 |
|
18 |
1 |
34.9 |
70.0 |
0.322 |
100 |
|
19 |
0.76 |
14.9 |
34.3 |
0.009 |
87 |
|
20 |
1 |
30.3 |
71.1 |
0.251 |
100 |
|
21 |
0.98 |
10.4 |
33.9 |
0.000 |
90 |
|
22 |
1 |
16.1 |
48.3 |
0.066 |
100 |
|
23 |
1 |
17.7 |
49.7 |
0.044 |
100 |
|
24 |
1 |
21.8 |
49.1 |
0.028 |
100 |
|
25 |
1 |
58.7 |
81.7 |
0.526 |
100 |
|
26 |
1 |
38.7 |
69.8 |
0.360 |
100 |
|
27 |
1 |
34.2 |
77.9 |
0.091 |
100 |
|
28 |
1 |
24.3 |
20.8 |
0.025 |
100 |
|
29 |
1 |
46.4 |
74.0 |
0.180 |
100 |
|
30 |
--- |
--- |
--- |
--- |
--- |
|
31 |
1 |
79.1 |
96.3 |
0.643 |
100 |
|
32 |
1 |
72.9 |
93.1 |
0.584 |
100 |
|
33 |
1 |
54.4 |
69.8 |
0.157 |
100 |

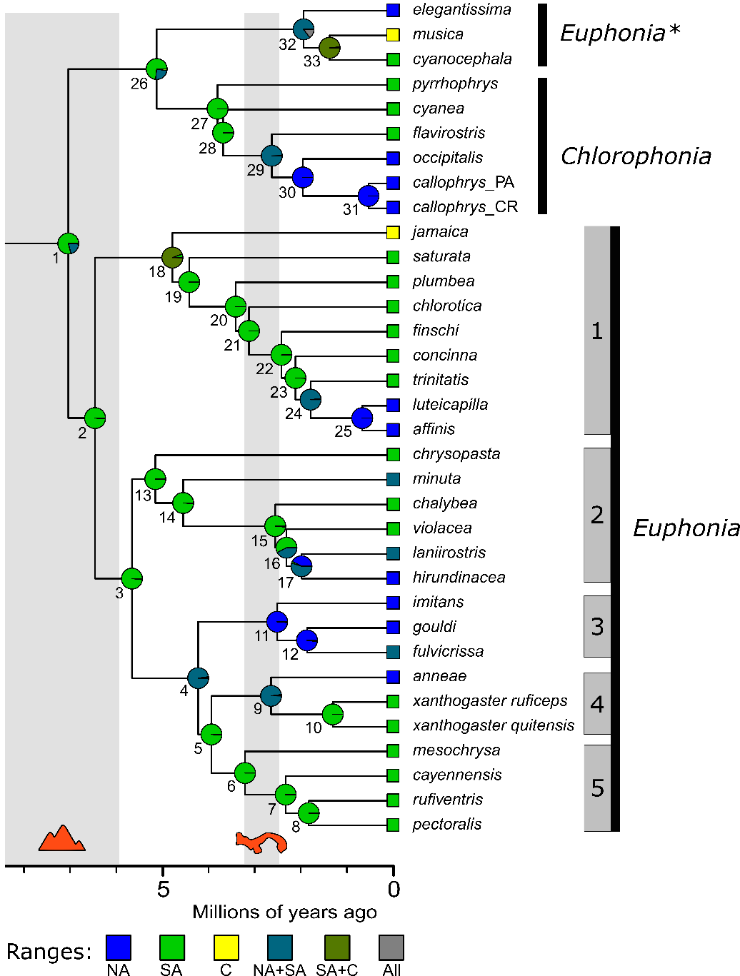
Figure
2. Maximum-likelihood phylogenies Euphoniinae generated from concatenated UCE
loci.
These trees are from Imfeld et al.’s Figure 1. Black nodes indicate bootstrap
support of 100%, and any node receiving <100% support has its value
reported. The topologies of these trees, especially the one inferred from all
UCE loci, are largely similar to the species tree topology presented above.
Triangles adjacent to the right tree indicate conflicting nodes between the
concatenated phylogenies.
Based
on the new phylogenetic information presented in Imfeld et al. (2020), we also
propose a new linear classification for the subfamily in the South American
checklist:
Cyanophonia
cyanocephala
Chlorophonia
cyanea
Chlorophonia
pyrrhophrys
Chlorophonia
flavirostris
Euphonia
saturata
Euphonia
plumbea
Euphonia
chlorotica
Euphonia
finschi
Euphonia
concinna
Euphonia
trinitatis
Euphonia
chrysopasta
Euphonia
minuta
Euphonia
chalybea
Euphonia
violacea
Euphonia
laniirostris
Euphonia
fulvicrissa
Euphonia
anneae
Euphonia
xanthogaster
Euphonia
mesochrysa
Euphonia
cayennensis
Euphonia
rufiventris
Euphonia
pectoralis
Due
to uncertainty in the phylogenetic placement of Euphonia jamaica and Euphonia
saturata, we have decided to collapse the node containing jamaica, saturata,
and the clade that contains the remaining species in ‘Euphonia clade I’
from Imfeld et al. (2020) into a polytomy. From this, jamaica should
precede saturata based on the northwest-to-southeast rule and this also
retains the current order of these two species in most classifications
(Clements et al. 2019). Similarly, we collapsed the node containing Chlorophonia
cyanea, C. pyrrhophrys, and, given this arrangement, cyanea
should precede pyrrhophrys following current the linear sequence of the
genus.
References:
Bonaparte, C.L. 1851. Note sur les Tangaras, leurs affinités, et descriptions d’espèces
nouvelles. Revue et Magasin de Zoologie Pure et Appliquée 2(3): 129-138.
Clements, J. F., T. S. Schulenberg, M. J. Iliff, S. M. Billerman, T. A.
Fredericks, B. L. Sullivan, and C. L. Wood. 2019. The eBird/Clements Checklist of Birds of the
World: v2019. https://www.birds.cornell.edu/clementschecklist/download/
Imfeld, T.S., F.K.
Barker, and R.T. Brumfield. 2020. Mitochondrial genomes and thousands of
ultraconserved elements resolve the taxonomy and historical biogeography of the
Euphonia and Chlorophonia finches (Passeriformes: Fringillidae).
The Auk: Ornithological Advances 137: 1-25.
Zuccon D., Prŷs-Jones
R., Rasmussen P.C., and Ericson P.G.P. 2012. The phylogenetic relationships and
generic limits of finches (Fringillidae). Molecular Phylogenetics and Evolution
62(2): 581-596.
Tyler S. Imfeld and
Nick A. Mason, May 2020
Note
on voting from Remsen:
let’s break into 2 parts: A. Cyanophonia. B. Linear sequence
Comments from
Areta: “A. NO. Looking
at the divergence times and phenotypic disparity within Euphonia, I feel
comfortable with leaving the blue-headed birds in Chlorophonia. Females
of Cyanophonia are very green, like Chlorophonias, and both male
and females have plumage patterns shared with other Cyanophonia (most
notably narrow foreheads, bluish crowns sometimes extending into semi-collars,
and petit bills). Additionally, songs of at least some species seem similar to
those of the blue-headed group in that they are jumbled series of quick notes,
and their low-pitched whistles are remarkably similar and unlike calls of Euphonia.
“Alternatively, if we split Cyanophonia from Chlorophonia,
then we would also be pressed hard to split Euphonia into more genera,
given the depth of the nodes and the consistency in plumage and vocalizations
of some of the groupings uncovered by Imfeld et al. (2020). This is the problem
I see when we have no more options to discuss on how to sort generic limits in
a comparative fashion. If there were sensible proposals on how to subdivide Euphonia
to match the level of phenotypic disparity and node depth of Cyanophonia,
I would be happy to reconsider my vote. For example, groups 1, 2 and 3+4+5
could be placed in different genera to match the split of Cyanophonia
from Chlorophonia. Unless this is done, separating Cyanophonia seems
to go against the generic concept being embraced for Euphonia.
‘The last, and worst, option would be to put all in Euphonia but
it would add too much diversity to the already diverse Euphonia, and
take the divergence times too deep into history for my taste.
“For these reasons, the simplest solution would be to put the
blue-headed Euphonia in Chlorophonia and not in a separate genus,
at least for the time being.
“B. YES.”
Comments from
Remsen: “A. NO, for
reasons outlined by Nacho. These are
great data, and a comprehensive phylogeny is a rare delicacy for us to deal
with. So, my “no” has nothing to do with
the underlying data but only the more subjective area of how to implement it in
classification. Based on node depth and
phenotypic heterogeneity, the implementation that I think is better is to just
transfer the Cyanophonia group into Chlorophonia. That
way the information content of the classification would reflect the phylogeny
in breaking the subfamily into two major lineages of roughly equivalent age,
thus facilitating comparisons between the two groups; their estimated node ages
(late Miocene, lining up nicely with similar estimates for the younger end of
taxon traditionally ranked as genera in birds).
That Chlorophonia was already
somewhat heterogeneous in pattern and size, and that true blue of the Cyanophonia group is also found in the plumages of 3 of 4
Chlorophonia species.
“As noted by Nacho, a case could also
be made for breaking Euphonia itself into
two genera, both of which would be older than Cyanophonia; in fact, from my viewpoint, if we recognize Cyanophonia, then we should also recognize taxonomically
at least two genera in the remaining euphonias, so I would vote no on the
proposal on that basis alone. As for
combining all into a single genus, I would object to that for many reasons,
including the new data in the paper that indicates a Miocene split.
“Tangentially, this group would be a
good one for using at least two subgenera to mark the major groups, assuming
there is a name available for Group 1.
“B. YES (spot
check of sequence indicates standard conventions followed, but I’ll check all.
Comments from Bonaccorso: “YES. Several arguments support
this proposal, as follows:
“1) "Cyanophonia" is
monophyletic and highly supported, and it is clearly diagnosable by plumage
(all with ‘cyano’ hoods).
“2) Although in terms of number
of changes, it seems easier to change three Euphonia* to Chlorophonia,
than three Euphonia* to "Cyanophonia", Chlorophonia
is a name that we all associate with green species (all very similar). If we
move Euphonia* to Chlorophonia, there will be three species of Chlorophonia
that are not green (not ‘chloro’), and Chlorophonia will no longer be
diagnosable based on plumage.
“3) I can't entirely agree with
the argument that if we resurrect Cyanophonia we will
have to split Euphonia. Splitting should not only be based on
synapomorphies and time of divergence. Taxonomic stability should also be
considered. Although some clades in Euphonia are diagnosable by plumage
and voice, splitting Euphonia would create a lot of unnecessary change
and confusion, especially for those who are not taxonomists.
“In conclusion, the resurrection of Cyanophonia allows for
naming three phylogenetically diagnosable units (Cyanophonia, Chlorophonia,
and Euphonia), having two morphologically diagnosable units (Cyanophonia
and Chlorophonia), and maintaining two well-established genera (Chlorophonia
and Euphonia), with minimum changes in the case of Euphonia.”
Comments
from Jaramillo:
“A. NO. I was thinking the same thing, having looked at the data, as what Nacho
wrote. It seems that you either separate Euphonia into multiple genera
or you include Cyanophonia in Chlorophonia. The least disruptive
seems to enlarge Chlorophonia.
“B.
YES.”
Comments from Zimmer: “A. NO, for reasons elucidated by Nacho and Van. I’m normally a proponent of more narrowly
defined genera, but, in this case, we have node depths that don’t support the
move, at least, not without further splitting of Euphonia, which is actually more supported by the data than is the
separation of Cyanophonia. I would definitely be opposed to lumping
everything into a very heterogeneous Euphonia,
but I would be entirely comfortable with having cyanocephala and the other two “Cyanophonia”
species placed within Chlorophonia. I can’t say that I agree completely with
Elisa’s Points #1 and #2 regarding Cyanophonia
being “clearly diagnosable” based upon their blue hoods, and that Chlorophonia would no longer be
diagnosable if the three Cyanophonia
species were included therein. If you
look at Chlorophonia, 4 of the 5
species have a blue hood (pyrrhophrys
and callophrys), a blue cap (occipitalis), or an extensively blue
nape (cyanea) of a very similar shade
of blue to that of Cyanophonia. Only the rather divergent flavirostris lacks blue on the
head/neck. Additionally, if you look at Cyanophonia, the females of ALL three
species are predominantly green, with yellowish median abdomens, and blue on
the crown & nape – VERY similar in pattern and color tone & saturation
to females of 2 of the aforementioned species (pyrrhophrys and callophrys)
of Chlorophonia, and differing from
females of the other 2 species (cyanea
and occipitalis) primarily in the
extent and distribution of blue on the head and nape. When both sexes are considered, I would argue
that neither powder blue on the head nor a predominantly green plumage is
sufficient to diagnose either Chlorophonia
or Cyanophonia from one another.
“B. YES, this linear sequence would appear to
conform to our conventions.”
Comments from Claramunt: “YES. Resurrecting Cyanophonia
makes perfect sense, resulting in a simple and descriptive classification. Three genera for three nicely delineated
groups: the typical euphonias (Euphonia), the blue-headed euphonias (Cyanophonia),
and the green euphonias (Chlorophonia) (note the matching Greek names
for color). I really do not understand
the arguments against this proposal. Merging Cyanophonia into Chlorophonia
would disrupt the diagnosability and homogeneity of two clearly delineated
groups. If the issue is the heterogeneity among typical euphonias, that should
be dealt with in a different proposal, not by killing this one.”
Comments from Stiles: “YES. Here, I agree with Santiago - the three
groups so defined are phenotypically and genetically distinctive. If this
requires subdividing Euphonia, then we can cross that bridge when we
come to it; and I note that the species remaining in Tangara show some
comparably deep divisions, if one were to further compromise stability for
consistency.”
Comments from Pacheco: “A. NO. After reading the various arguments, I think that the
results of phylogeny point to the inclusion of "Cyanophonia"
in Chlorophonia as the most appropriate decision. B. YES.”