Proposal (864) to South American Classification Committee
Elevate Podiceps occipitalis juninensis to species rank
Background. The form of
Silvery Grebe from the Central Andes was originally named as a subspecies, and
indeed bears close resemblance to the nominate race of Patagonia, except when
the latter is in its striking breeding plumage. Lacking a distinct non-breeding
plumage, juninensis is actually more
similar to Junín Grebe Podiceps
taczanowskii in all adult plumages except for its smaller overall size but
relatively longer wings, to the extent that specimens can be difficult to
distinguish even in the hand (Peters and Griswold 1943). Until recently, juninensis has generally been treated as
a subspecies, although Chubb (1919) treated it as Podiceps juninensis,
without further comment but citing Ogilvie-Grant (1898) who, however, had
clearly captioned his account, which followed that of P. occipitalis, “Subsp. a.
Podicepes juninensis”.
New Information. More recently, some authors have suggested that juninensis may be better treated as a full species, and del Hoyo
and Collar (2014) split juninensis on
the grounds of morphology, non-migratory behavior, lack of distinct breeding
plumage, and higher-pitched voice. In a phylogeny of cytb and COI (see screenshot below), Ogawa et al. (2015) found that juninensis, taczanowskii, and occipitalis
formed a clade, but that the two samples of juninensis
(which were not reciprocally monophyletic) were basal to the clade containing taczanowskii, with the single sample of
nominate occipitalis embedded within taczanowskii. Ogawa et al. (2015)
suggested that the lack of reciprocal monophyly between the two juninensis samples may be due to the
isolated Colombian Andes population possibly being a genetically distinct
undescribed taxon. In any case, according to this phylogeny, juninensis is less closely related to taczanowskii and occipitalis than the latter two are to each other, and the
estimated age of divergence between juninensis
and taczanowskii + occipitalis is a little more than 1.5
myr.
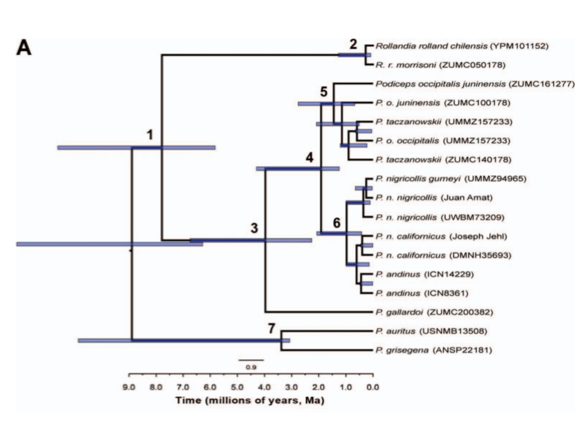
Fig. 2A of Ogawa et al. (2015).
Conclusions. The case of P. occipitalis juninensis seems a
somewhat analogous situation to Andean Teal Anas
andium and Puna Teal Spatula puna,
which are now considered full species by SACC (Remsen et al. 2020), and a
stronger case for species status than the Colombian Grebe P. andinus, which is embedded within Eared Grebe P. nigricollis as currently recognized.
Proposed Changes. A YES vote for Part A of this proposal (strongly recommended) would be
for the treatment of juninensis as a
full species, Podiceps juninensis.
As for English names (if A passes), ‘Northern
Silvery Grebe’ was used for P. juninensis
and ‘Southern Silvery Grebe’ for P.
occipitalis by del Hoyo and Collar (2014); they are apt and have gained
currently through HBW/BLI and e.g. Guevara et al. (2016), but they are a bit
long and bland. The group names used by Clements et al. (2019) are ‘Andean’ and
‘Patagonian’, respectively; if used in combination with ‘Silvery’, ‘Patagonian
Silvery Grebe’ is overlong with 9 syllables. Only two grebes are widespread in
the Andes, and only one is widespread yet a breeding near-endemic to Patagonia
(though Great Grebe Podiceps major comes
close). A hybrid option would be ‘Andean Grebe’ and ‘Silvery Grebe’, which has
the advantage of retaining the name of the most widespread form.
Thus, if voting yes for A please vote on Part
B for either ‘Northern Silvery Grebe’ and ‘Southern Silvery Grebe’; ‘Andean
Grebe’ and ‘Patagonian Grebe’, ‘Andean Grebe’ and ‘Silvery Grebe’, or a
suggested permutation of these.
References.
Chubb, C. (1919). Notes on collections of
birds in the British Museum, from Ecuador, Peru, Bolivia, and Argentina. Part
II. Podicipediformes―Accipitriformes. Ibis 1919: 256–290.
Clements, J. F., T. S. Schulenberg, M. J.
Iliff, S. M. Billerman, T. A. Fredericks, B. L. Sullivan, and C. L. Wood
(2019). The eBird/Clements Checklist of Birds of the World: v2019. Downloaded
from https://www.birds.cornell.edu/clementschecklist/download
del Hoyo., J., and N. J. Collar (2016). HBW
and BirdLife International Illustrated Checklist of the Birds of the World.
Volume 2: Passerines. Lynx Edicions, Barcelona.
Guevara, E. A., T. Santander G., A. Soria, and
P.-Y. Henry (2016). Status of the Northern Silvery Grebe Podiceps juninensis in the northern Andes: recent changes in
distribution, population trends and conservation needs. Bird Conservation
International 26: 466–475.
Ogawa, L. M., P. C. Pulgarin, D. A. Vance, J.
Fjeldså, and M. van Tuinen (2015). Opposing demographic histories reveal rapid
evolution in grebes (Aves: Podicipedidae). The Auk: Ornithological Advances
132: 771–786.
Ogilvie-Grant, W. R. (1898). Catalogue of the
Birds in the British Museum. Vol. 26.
Steganopodes (Cormorants, Gannets, Frigate-Birds, Tropic-Birds, and
Pelicans), Pygopodes (Divers and Grebes), Alcae (Auks), and Impennes
(Penguins). Trustees, London.
Peters, J. L., and J. A. Griswold, Jr. 1943.
Birds of the Harvard Peruvian Expedition. Bulletin of the Museum of Comparative
Zoology 92: 280–327.
Remsen, J. V., Jr., J. I. Areta, C. D. Cadena,
S. Claramunt, A. Jaramillo, J. F. Pacheco, J. Perez Emán, M. B. Robbins, F. G.
Stiles, D. F. Stotz, and K. J. Zimmer (Version 11 February 2020). A
Classification of the Bird Species of South America. American Ornithological
Society. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm
Pamela C. Rasmussen,
July 2020
Note on
Remsen for voting: Unless one of the proposed English names gets a 2/3 majority
from English name subcommittee, I’ll convert the results into a separate
proposal that uses the winner of this vote as the starting point.
Comments
from Robbins: “YES. To help evaluate differences in breeding plumage that Pam
mentions, I'm pasting in a few eBird checklists that have unequivocal birds in
breeding plumage of each occipitalis taxon:
https://nam04.safelinks.protection.outlook.com/?url=https%3A%2F%2Febird.org%2Fchecklist%2FS53665186&data=02%7C01%7Cnajames%40lsu.edu%7C0be9fe96f39849361d5808d82cb33ded%7C2d4dad3f50ae47d983a09ae2b1f466f8%7C0%7C0%7C637308495634499516&sdata=6R4NxdoQRxrfS7%2BrYz2jJc2kiSOU2E6SAeOGySkqpxM%3D&reserved=0
https://nam04.safelinks.protection.outlook.com/?url=https%3A%2F%2Febird.org%2Fchecklist%2FS21087057&data=02%7C01%7Cnajames%40lsu.edu%7C0be9fe96f39849361d5808d82cb33ded%7C2d4dad3f50ae47d983a09ae2b1f466f8%7C0%7C0%7C637308495634499516&sdata=wQPsnycmZqoHkAg6HxWoeKEHgSONWV2rX2G4KhRkD7Y%3D&reserved=0
https://nam04.safelinks.protection.outlook.com/?url=https%3A%2F%2Febird.org%2Fchecklist%2FS27026845&data=02%7C01%7Cnajames%40lsu.edu%7C0be9fe96f39849361d5808d82cb33ded%7C2d4dad3f50ae47d983a09ae2b1f466f8%7C0%7C0%7C637308495634499516&sdata=WqHSsDBIw8M5ekx0vkRj6EjesMSRrHCv400LXYNqNfo%3D&reserved=0
https://nam04.safelinks.protection.outlook.com/?url=https%3A%2F%2Febird.org%2Fchecklist%2FS65088774&data=02%7C01%7Cnajames%40lsu.edu%7C0be9fe96f39849361d5808d82cb33ded%7C2d4dad3f50ae47d983a09ae2b1f466f8%7C0%7C0%7C637308495634499516&sdata=XQB8bob9XAr662Hr7y%2FQOelM2OfoMIU6njscz6cZ0M4%3D&reserved=0
Comments from Areta: “NO. The issue with these grebes is not
so easy from my perspective, and while there are extremes of (mostly)
geographically structured morphological variation that have been usually
recognized as juninensis, taczanowskii, and occipitalis, I
feel that more information is needed to recognize juninensis as a full species. The situation might be more fluid
than what current studies show, and we may be jumping the gun by simplifying
the case.
“It is worth noting
that in several paragraphs Ogawa et al. 2015 refer to "incipient
species" and not to "full species" when mentioning taczanowskii and andinus. The first two paragraphs of the discussion show the
careful approach of the authors when dealing with taxonomic treatment of these
forms. And the last paragraph of Ogawa et al. 2015 (p.781) indicates that
"we conclude that the now extinct P. andinus represented a newly
established lineage and incipient species among Podicipedidae. Furthermore, and
consistent with a tendency for rapid speciation in grebes, P. andinus may represent one of several incipient species, as is
indicated by DNA barcode data on P.
taczanowskii (Junin Grebe; this study) and the Aechmophorus occidentalis–A.
clarkii (Western Grebe– Clark’s Grebe) complex"
“First of all, Thomas
Valqui first and myself later found many individuals at Lake Junin (where taczanowskii and juninensis coexist) that were difficult to identify to species.
This may suggest that there is ongoing hybridization between them (perhaps
triggered by the recent population declines in taczanowskii? for which see Valqui 1994. Or perhaps this was always
the case?). I am unaware of published information in this regard.
“Second, the
phylogenetic relationships shown in the Cyt-b+COI tree differ from the control
region (Figure 3 in Ogawa et al. 2015). If we focus on the Cyt-b+COI tree, then
both taczanowskii and juninensis would be non-monophyletic:
with one juninensis (from Lake Junin) being more closely related
to occipitalis+taczanowskii than to the other juninensis
(from Lagunillas); similarly, the clade of occipitalis+taczanwoskii shows that the single
sample of occipitalis is more closely
related to a taczanowskii than to the
other sample of taczanowskii. In
looking at the control region inset (Clade I), the Colombian birds seem more
distinct than juninensis, occipitalis and taczanowskii from Peru and Argentina, while the relationships among
the latter three differ from those in the Cyt-b+COI tree: there is a polytomy
involving occipitalis (Argentina), taczanowskii (Junin) and a clade
including one taczanowskii (Junin)
and one juninensis (Peru). Thus, for
different reasons, taczanowskii and juninensis are not monophyletic in both
datasets. The only solid conclusion that I can draw from Ogawa et al. (2015) is
that genetic differentiation between juninensis,
taczanowskii and occipitalis is reduced, and that more samples and further genetic
analyses are needed to understand how genetic variation parses out among. I do
not think that paper provides compelling evidence for a split (and the paper
itself does not claim this).
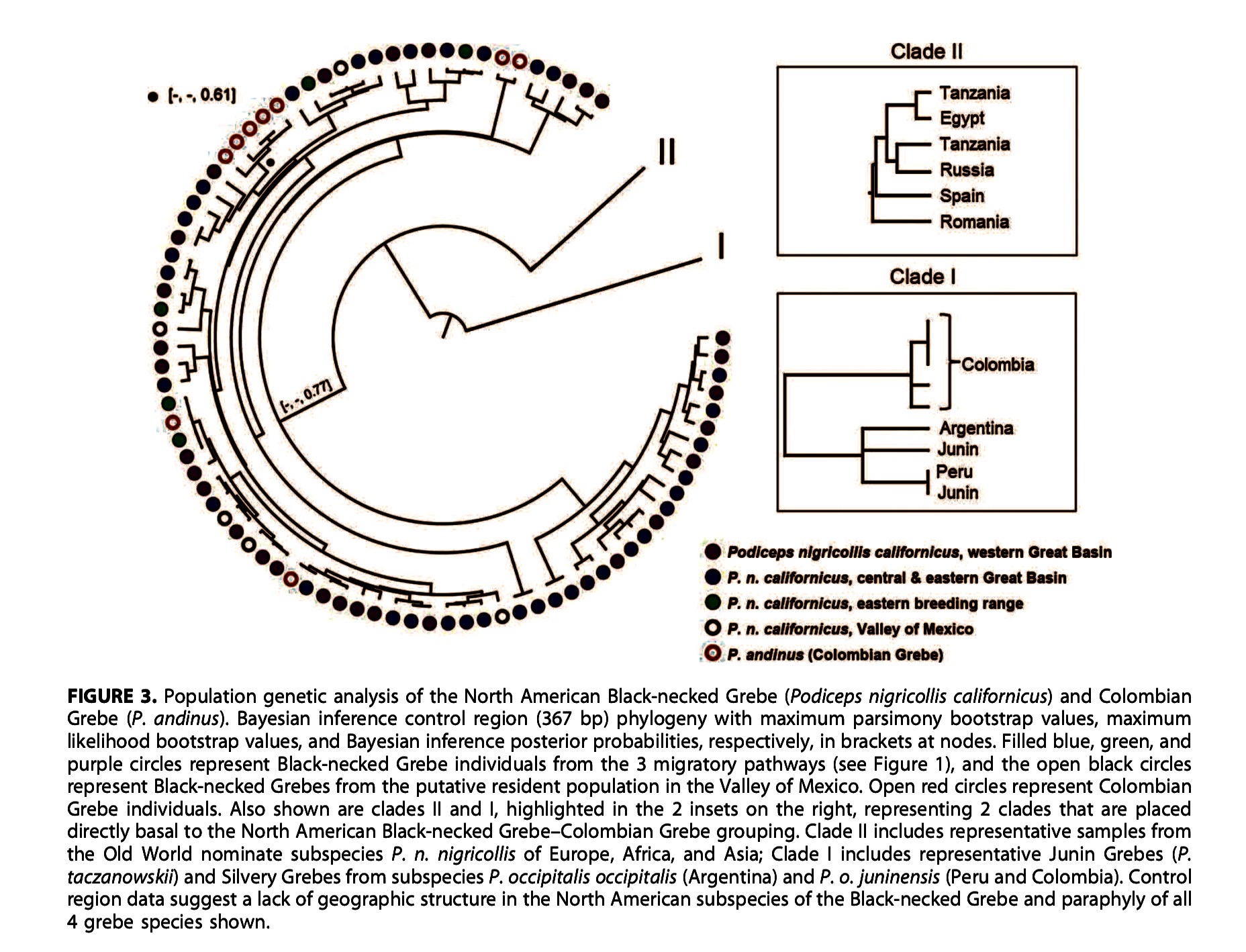
“To conclude, I find
the genetic data presented in Ogawa et al. (2015) as insufficient evidence to
elevate juninensis to species. A
stronger case for the split could possibly be made by integrating vocalizations
(there are no published analyses, not even spectrograms), differences in
plumage patterns (indeed, taczanowskii
is more similar to juninensis than to
occipitalis) and geographic
distributions while assessing for the existence of intermediates in a
comparative phylogenetic framework (this is missing, as no one has really
looked into this). The plumage differences between juninensis/taczanowskii
and occipitalis seem clear, but are
not as marked as those between undisputed species. As Alvaro mentioned for
Chile, occipitalis and juninensis also overlap to some extent
in the Puna of NW Argentina, but no one (to my knowledge) has studied the
situation in detail. Until solid genetic and natural history data is properly
analyzed, I prefer to err on the side of caution by leaving juninensis as an arguably quite
diagnostic subspecies of occipitalis.
In the long run, the evidence may show that occipitalis
and juninensis are different species,
but this evidence should be gathered systematically, carefully analyzed and
published. At present, I see many gaps, conflicting data and key unanswered
questions to make this decision.
“See: Valqui,
T. (1994) The extinction of the Junin Flightless Grebe? Cotinga 1:
42–44.”
Comments
from Jaramillo:
“YES. Additional
information on these from Chile: On several occasions we have seen occipitalis
in flocks of juninensis in Lake Chungara up by the Peru/Bolivian
borders. Steve Howell has also found them up there in flocks of juninensis.
This is in the October-November time frame, so breeding season. Note that there
is a well-documented record of occipitalis in southern Peru. Apart from
the golden plumes, look at the dark throat and chin which juninensis
will not show. This is a September record, so pre-breeding season for occipitalis. https://nam04.safelinks.protection.outlook.com/?url=https%3A%2F%2Febird.org%2Fchecklist%2FS48454501&data=02%7C01%7Cnajames%40lsu.edu%7C47c87a67084743b1da2008d82cdfaea8%7C2d4dad3f50ae47d983a09ae2b1f466f8%7C0%7C1%7C637308686527250172&sdata=MV6m%2FzrQxA50OX1ezO6LZWWnfRMMxopJ2yCC4fzyxVI%3D&reserved=0
“There is no way to assess what is going on, other that there is
sometimes sympatry during the early part of the breeding season (on Chungara we
see nests of juninensis in Oct-Nov). The occipitalis may be
migrants that will leave, I don't know? I assume that is the case. There is no
evidence of hybridization that we know of. But do keep in mind that occipitalis,
unlike juninensis, is highly migratory. Migration and distinct change
from a breeding to a non-breeding plumage is of interest, versus juninensis
that looks about the same year round and is resident.”
Comments from Claramunt: “NO. Differences in facial
patterns are very suggestive, I admit I was tempted to vote YES, but we need
some minimal analysis of the geographic distribution of the character to
evaluate the potential existence of intermediate populations. And then, the Ogawa
et al. paper shows a very complicated picture of mitochondrial relationships
with no clear genetic clades coinciding with taxonomy nor with phenotypic
similarity, suggesting no clear separation of species-level lineages, including
taczanowskii. I could not find the DNA sequences in GenBank, so no
chance of doing some further analyses (strange, I think The Auk requires
publication of sequences). Rather than splitting juninensis, these
results rise the question of whether taczanowskii should be considered a
separate species (cf. also Nacho’s comment).”
Additional comments from Jaramillo: “I was
looking through where the closest points of contact may be between these two
forms, and seeing if I could come up with some imagery. An interesting record I found is this one from
Jujuy in January with both forms together. Apart from the throat coloration differences,
and the golden plumes versus silvery the crown/nape contrast is another
difference. The form juninensis
has a darker crown, not as contrasting as on occipitalis. They also look
different from behind. In any case, I thought this record of sympatry would be
interesting:
Additional comments from Claramunt: “Alvaro,
I hear you. Phenotypically, they seem
two different species, in my mind. But the genetic data suggest a different
scenario. The facial pattern may be the
result of a single (or just minimal) genetic difference in a background of
genomic uniformity (or at least not diverging).
The genetic basis of the trait may not
allow for intermediate forms and interbreeding may be invisible. Then the presence of both phenotypes
in a single population can be interpreted as a local polymorphism and a
confirmation that the two phenotypes behave, socially, as a single species. The point is that, without additional data, we
cannot interpret co-occurrence as evidence of reproductive isolation.
“If they are breeding sympatrically, observing whether pairs sort
out by phenotype or whether there are phenotypic intermediates would be very
informative.”
Comments
solicited from Jon Fjeldså: “I am glad to see that you take up the ranking of these
grebes, as I was quite irritated to see P. occipitalis and juninensis
being split in The HBW/BirdLife illustrated checklist. However, I can probably blame myself, since
nearly 40 years ago I myself published some evidence for splitting them (see
Fjeldså J 1982. Some behaviour patterns of four closely related grebes, Podiceps
nigricollis, P. gallardoi, P. occipitalis
and P. taczanowskii, with reflections on phylogeny and adaptive
aspects of the evolution of displays. Dansk Orn. Foren. Tidsskr. 76: 37-68. Here, I described some differences in displays
and calls between silvery grebes in Peru and southern Patagonia. However, these populations are far apart, and
I really don't know how much variation there could be in behavior within the
large intervening area. In fact, there seems to be a good deal of flexibility
in grebe behavior, in whether they use face-to-face dancing or parallel rushes
or 'barging'.
“Since
then, I discovered perfectly intermediate specimens (between occipitalis
and juninensis) in museum material from around Coquimbo in Chile. This
suggested a rather narrow zone of genetic mixing in the zone of contact between
southern and northern morphotypes, but in fact there are also indications of
geneflow over deeper time since juninensis birds from northern Chile,
Bolivia and southern Peru (Puno, Arequipa) generally have somewhat
golden/brass-like luster to the ear plumes (but white throats), in contrast to
dull greyish-brown ear-plumes in all populations from Junín and Ancash through
Ecuador and Colombia. So clearly there is a rather gradual transition in
morphology between occipitalis and juninensis (but no detectable
difference between Junín-Ancash and the northern Andes).
“The
result by Ogawa et al. was surprising (note that I was co-author here), because
of the genetic break between specimens from the south+Junín+Ancash and the
northern Andes. So there is a discordance between morphological and molecular
signals here. I think many more genetic samples are needed (with multiple
markers for telling apart effects of gene flow and ancestral polymorphism)
before we can tell whether there is a basis for splitting up the Silvery Grebe.
With present techniques, it should be quite straight-forward to obtain adequate
genetic data (broad geographical sampling) from toepads of museum specimens,
with no need for collecting fresh samples.
“P.
taczanowskii is another issue. Although placed next to an occipitalis
sample in the Fig. by Ogawa et al., there is no doubt from my old work in Junín
that this population, which probably originated during a period of glacial
isolation in the Junín Basin, is reproductively isolated from juninensis.
P. juninensis and taczanowskii may sometimes display together,
but using rituals that have nothing to do with pair formation (but rather a
sign of mild/ritualized aggression), and in many contexts, for instance during
foraging, they decidedly avoid one another. They are clearly aware that they are
different. I am rather skeptical about
claims of intergradation by the two species in lake Junín. On the contrary, my morpho-data (in several
publications) indicate divergence (ecological character displacement), but I
have seen one bird that could be an hybrid (but casual hybrids are also known
between occipitalis and gallardoi, and between other well-marked
grebe species). To check if there could have been a recent breakdown in the
genetic integrity of taczanowskii I checked photos available on Google
(assuming that most of these have been uploaded in recent years). One bird
labelled as taczanowskii was a clear occipitalis, 3 were
difficult to identify (because of angle of view, or resolution), but 71 were
clearly taczanowskii. Therefore, I don't see any evidence of
hybridization but rather think that some people have difficulties with
identifying the species. Again: more genetic data are needed. Ogawa et al.
provided some indication that isolated local populations of grebes can rapidly
diverge and evolve their own integrity, although they are nested within
widespread species in phylogenies using few genetic markers.
“There
is no reference to any of my old papers (1980s) on Peruvian grebes in the reference
list to Proposal 864 (reprints of these should be deposited in several American
museums, and at least they are summarized in my 2004 book 'The Grebes' from
Oxford Univ. Press).
“My
conclusion: Keep occipitalis and juninensis as one species and
maintain taczanowskii (as well as andinus, based on the arguments
provided by Ogawa et al.) (a separate question is then whether Podiceps nigricollis
should be split up with separate species in the Nearctic and in the Old World
(and maybe a third species in South Africa).
“I
hope this is useful for your decision.”
Comments
from Bonaccorso:
“NO. The case is complicated in so many dimensions
(plumage, vocalizations, natural history, distributions) and so unclear
regarding phylogenetic evidence (e.g. cytb and COI vs. control region) that I
think that elevating this taxon to species requires a paper on its own. As said
before, a systematic revision of all qualitative and quantitative evidence and,
probably, more genetic information will be needed to make an informed decision.”
Comments from Pacheco: “NO. Further studies may show
that occipitalis and juninensis are different species, but this
evidence still needs to be collected in the key areas and analyzed in an
integrated manner. As highlighted by Nacho, the article by Ogawa et al. does
not provide convincing evidence for a division.”
Comments from Zimmer: “NO. I was on the fence with this one, mainly
due to occipitalis being migratory
and having distinct breeding and non-breeding plumages, in contrast to juninensis. However, after considering comments from
several other committee members, and, in particular, Jon Fjeldså, I am
persuaded that we simply don’t know enough about what is going on to justify
splitting at this time.”
Additional
comments from Jaramillo: “I realize that I am against the tide here, but that is
fine. I think longer term with more data we will sort this one out as two
species. There seem to be some unusual aspects in grebes that I certainly do
not understand. Aechmophorus grebes breed sympatrically, sometimes
hybridize but actually tend to mate assortatively from what I understand. Vocal
differences are there, but minor really, plumage and bill color differences
consistent but minor. Podiceps nigricollis has these relatively large
genetic differences between populations (New World and Old World separated 1
million years ago), yet little morphological, or at least plumage differences.
In the New World we have three examples at least of flightlessness developing,
and for the most part the plumage of these birds has remained (at least 2 of 3)
quite similar if not nearly identical to the flying sister species. In all
cases the sister is sympatric with the flightless population. It is a mystery
to me how this happens, was it sympatric speciation? If so, how come the
plumages are so conserved? Then we have the rather spectacular array of
face/neck ornamentation in this group, and often a molt into a dull
non-breeding plumage. Surely these ornaments are of some use, some purpose and
some use in pair formation.
“In any case, I considered Santiago’s points
and I do not think that this is what is going on. I do not think that this is a
morph, or at least a situation where hybrids may go undetected due to a genetic
situation where intermediates would not be expressed. As I looked at more
photos, there does seem to be a zone where you can find intermediates, and
Fjeldså noted this as well. I am not sure about his interpretation of a wide
zone of gene flow. But there is a contact zone, and certainly some hybridization.
As such perhaps some would suggest that this be studied in more context, and
that is reasonable. But at this point the differences in ecology, display
features (breeding plumes), resident vs migrant situation, and seemingly little
or a narrow zone of interbreeding. I am sticking with my YES vote to split
these two.”
Comments from Remsen: “NO. Alvaro may be correct, but this is a
complicated case that needs more research before we make a change, as noted by
several comments above, including Jon’s, whose input is much appreciated.”