Proposal (868) to South American Classification Committee
Treat
Lepidocolaptes layardi as a subspecies of L. fuscicapillus
Effect on South American CL: If adopted, this proposal would lump two taxa of the genus Lepidocolaptes
presently considered species and would thus recognize the polytypic species L.
fuscicapillus (with nominate and layardi subspecies).
Background: Proposal 620 (2013) proposed the elevation to species rank of all taxa included in
the species previously named Lineated Woodcreeper (Lepidocolaptes
albolineatus). This proposal was adopted in 2014 and has subsequently been
taken over in several worldwide taxonomic lists. (For the background until that
date, see the proposal.)
New information:
1. Proposal 620 states in the background info: “taxa fuscicapillus
and layardi being very distinct vocally, hence suggesting that the
polytypic L. albolineatus may include more than a single species
(Marantz et al. 2003).” However, what is described in this reference for fuscicapillus
is in fact the voice of the (then undescribed) fatimalimae. On the
contrary, they describe the voice of madeirae (=present fuscicapillus)
and layardi as being the same.
2. Proposal 620 states in the new information: “Vocally, these five
molecular clades/taxa have also proved to be very distinct, further supporting
their treatment as independent species (Rodrigues et al. 2013).” However, the
‘proof’ given in Rodrigues et al. (2013) for vocal difference between fuscicapillus
and layardi is nowhere to be found in the text (this publication is in
fact about the description of the new taxon fatimalimae, and only the
vocal difference of this taxon vs. all others is detailed. There is, however, a
single picture of ‘representative sonograms’ that suggests a difference in
voice between fuscicapillus and layardi (see also further) and a
Table 1 in Supplementary Information. I am not sure if this supplementary
information is still available on-line somewhere, for which I copy it here:
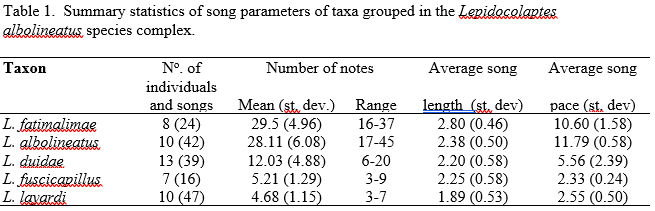
From this Table, it is clear that the measured sound
parameters show no significant differences for the taxa fuscicapillus
and layardi (at most a slight difference in song duration with
considerable overlap).
3. Boesman (2016) made a brief analysis of the voice of the Lineated
Woodcreeper complex. As no clear vocal differences could be found between fuscicapillus
and layardi, the specific sonogram depicted in Rodrigues et al. (2013)
was traced back to the original recording. When depicting with appropriate
amplitude levels, the note shape appeared to be quite different and more in
line with other sonograms of both fuscicapillus and layardi (with
note shapes slightly dependent on excitement level of the bird).
4. Independently, Minns (2016) also catalogued all existing xeno-canto
recordings. He states: “The song of layardi is similar to that of its
neighbour fuscicapillus and I was not able to readily distinguish
between them.”
From the above, I believe there is at present no
indication at all that voice of fuscicapillus and layardi shows
anything more than some minor differences.
Rodrigues et al. (2013) also supplied genetic
information (albeit, similar to the vocal information, also in a very condensed
way without providing much detail). From the figure 1 it appears that fuscicapillus
and layardi are the two (sister) taxa which have the smallest genetic
divergence.
Analysis and Recommendation: We have thus here two taxa which at most show some minor morphological
and vocal differences, and which have a considerable mtDNA divergence, which,
however, is the smallest compared to other members of the complex (the latter
additionally DO show significant vocal differences among them as confirmed in
Boesman (2016) and Minns (2016) !).
From the comments of the SACC members on proposal 620,
I deduce that the vocal difference was the main argument to split Lepidocolaptes
albolineatus into four species. With the new information presented here,
this vocal difference is no longer present for the taxa fuscicapillus
and layardi.
Whether the genetic divergence in itself is sufficient
argument to maintain both taxa as full species obviously depends on which
criteria are used. Accepting BSC species level purely on a moderate mtDNA
divergence for two populations separated by a geographical barrier seems at
least rather adventurous and may open the discussion what to do with many other
similar cases.
It is clear that the Tapajos (and Tele Pires) river is
an important geographical barrier, which has led to isolation and speciation in
quite some cases. This in itself however can’t be an argument. After all, Duida
Woodcreeper L. duidae also occurs on both sides of the Rio Negro, an
equally important geographical barrier while at the other hand, it is only
separated from L. albolineatus by the ‘small’ Rio Branco.
Therefore, I propose to reconsider the decision taken
in 2013, would recommend lumping the two taxa and recognize the polytypic
species L. fuscicapillus (with nominate and layardi subspecies).
As for the English name, del Hoyo and Collar (2016)
lump both taxa as a single species and have given it the name Dusky-capped
Woodcreeper (reflecting its scientific name).
Literature cited
Boesman,
P. (2016). Notes on the vocalizations of Lineated Woodcreeper (Lepidocolaptes
albolineatus). HBW Alive Ornithological Note 84. In: Handbook of the Birds
of the World Alive. Lynx Edicions, Barcelona. https://static.birdsoftheworld.org/on84_lineated_woodcreeper.pdf DOI: 10.2173/bow-on.100084
del
Hoyo, J., Collar, N.J. (2016). HBW and BirdLife
International illustrated checklist of the birds of the world. Volume 2:
Passerines. Lynx Edicions, Barcelona.
Rodrigues,
E. B., Aleixo, A., Whittaker, A., and Naka, L. N. (2013). Molecular systematics and taxonomic revision
of the Lineated Woodcreeper complex (Lepidocolaptes albolineatus:
Dendrocolaptidae), with description of a new species from southwestern
Amazonia. In J. del Hoyo, A. Elliott, J.
Sargatal & D. Christie (Eds.), Handbook of the Birds of the World. Special Volume: New Species and Global Index,
pp.248-252. Lynx Edicions, Barcelona.
Marantz,
C., A. Aleixo, L. R. Bevier and M. A. Patten (2003). Family Dendrocolaptidae
(Woodcreepers). Pp. 358-447 in: del Hoyo, J., A. Elliott, and D. A. Christie
(eds.) (2003). Handbook of the Birds of the World. Volume 8: Broadbills to
Tapaculos. Lynx Edicions, Barcelona.
Minns,
J. (2016) Delineating the Lepidocolaptes albolineatus complex. https://www.xeno-canto.org/article/198
Peter
Boesman, July 2020.
Comments
from Remsen:
“YES (depending on comments by others). The
evidence above convinces me that the evidence for the split was over-stated and
not independently analyzed by us as thoroughly as it should have been. With respect to genetic differences, when
restricted to a few neutral loci from a few individuals within taxa of a
monophyletic group, in my viewpoint they are irrelevant to taxon rank.”
Comments from Whitney: “I appreciate with
this well-explained argument of Peter’s, and certainly concur that layardi
is minimally differentiated (if at all) from fuscicapillus. This case is similar to those of other
"HBW woodcreeper" cases (Campylorhamphus, Dendrocolaptes) in
which sample sizes of data used in diagnostic analyses, phenotypic and genetic,
were inadequate (regardless of the conclusions drawn from them).”
Comments
from Claramunt:
“YES. The previous splitting of this species
was done without critical evidence.”
Comments from Robbins: “YES, given Peter Boesman's assessment.”
Comments from Stiles: “YES. Strong vocal similarity and weak genetic
separation without evidence of reciprocal monophyly combine to make its species
status unsustainable.”
Comments from Areta: “YES. The Lepidocolaptes seem to be quite geographically variable in plumage,
but these differences are not paralleled by differences in vocalizations. This
provides an interesting comparison to the case of Lepidocolaptes squamatus/falcinellus in relation to the level of
isolation and the roles of rivers in promoting/maintaining differentiation.
Comments solicited from Jorge Pérez-Éman:
“This
proposal aims to lump Lepidocolaptes
fuscicapillus and L. layardi into
a single polytypic species (L.
fuscicapillus), partially reverting the decision to split L. albolineatus into five species,
including recently described L.
fatimalimae (Rodrigues et al. (2013), Proposal 620). In this proposal,
Boesman argues that data to consider layardi
different from fuscicapillus is
unavailable, and makes the following points:
“1. Early
statements about the large differences between fuscicapillus and layardi
vocalizations were influenced by the known distribution of both taxa by that
time (Marantz et al. 2003). The currently known distribution for each taxon in
the entire L. albolineatus complex is
now better understood by the work of Rodrigues et al. (2013) and, as pointed
out by Boesman, such differences referred to fatimalimae (and not fuscicapillus)
and layardi.
“2.
Rodrigues et al. (2013) provided a molecular phylogeny and stated that the five
monophyletic lineages obtained from this hypothesis proved to be very distinct
vocally. However, Boesman rightly points out that Rodrigues et al. (2013)
focused only on the differences between fatimalimae
and the other taxa, and no detailed analyses were done on the other four taxa (albolineatus, duidae, fuscicapillus and
layardi). He goes further and
indicates that no clear vocal differences were found between fuscicapillus and layardi, and back this up with some general and qualitative
comparative vocal analyses done within this complex (Boesman 2016, Minns 2016).
“I totally
agree with Boesman in relation to these general comments. Rodrigues et al.
(2013) focused on the description of fatimalimae
and consequently, a proposal to split L.
albolineatus into five taxa was not really well supported by their data and
analyses. I think the authors did a great job to better understand the
diversity and distribution of taxa within this species complex, but I don´t
know if by omission or by constraints provided by HBW publishers, there is not
enough information to evaluate the validity of such split. As early reviewers
of Proposal 620 indicated, I was really frustrated trying to find details
associated to detailed information on skin vouchers, recordings and tissues
used in the study. First, the map included in the article is not clear and
details are impossible to see and, second, I am still looking for the
Supplementary Information (besides the Table S1 provided by Boesman). This
information should be readily available to readers/reviewers and it is
particularly relevant when taxonomic changes are proposed. In particular, I
missed a more complete treatment of the morphological variation (although some
of the Zimmer’s [1934] concerns on morphological variation were partly solved
by description of fatimalimae),
including patterns in contact zones among different taxa. Same goes for vocal
and molecular variation. For example, available information for both
morphological/molecular (Rodrigues et al. 2013) and vocal data (Minns 2016)
suggest sympatry in Bolivia, but hybridization has not yet been assessed. A
more thorough specimen evaluation in this region would have provided with a
stronger support to some of their proposed splits by confirming sympatry
without hybridization. Similarly, what are the patterns on both sides of the
Madeira, Tapajos or Teles Pires rivers? These areas have been known to show
complex patterns of phenotypic/molecular variation in other taxa and clear
definition of species/taxon limits should take this into account.
“Boesman´s
criticism on vocalizations analyses done by Rodrigues et al.(2013) are worth to
expand here. The description of L.
fatimalimae vocalizations and their differences to all other taxa within
the L. albolineatus complex are very
detailed and complete. However, there is no similar analysis done with other
taxa vocalizations and the only results shown are included both in Table S1 and
a figure with spectrograms of songs for each taxon. I am providing plots for each
of the vocal characters included in Table S1 to better visualize patterns
(Figure 1).
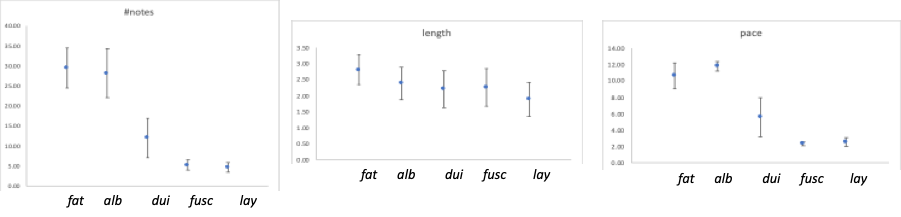
“Figure 1.
Vocal characters used to compare taxa within the Lepidocolaptes albolineatus species complex (Rodrigues et al.
2013). Number of notes, song length (seconds) and pace (number of notes per
second) data are extracted from Table 1 included in the Supplementary
Information of the HBW Special Volume. Dots represent means and error bars one
standard deviation. Taxa abbreviations: fat
= fatimalimae, alb = albolineatus, dui = duidae, fusc = fuscicapillus, lay = layardi).
“As you can
see, and clearly pointed out by Boesman, these data show no differences between
fatimalimae and albolineatus, or between fuscicapillus
and layardi vocalizations for any of
the three variables. It is also interesting that the first three taxa show a
larger variation in these characters than the last two and, given this
variation, duidae slightly overlaps
with both fuscicapillus and layardi. In fact, songs of these three
taxa were found to be similar by Marantz et al. (2003) describing them as
“slowing, descending series of clear whistles” differing mainly in number of
notes and pace. Moreover, characters such as note shape, frequency, inter-note
intervals and the acceleration/deceleration pattern, useful in the comparisons
with fatimalimae, were not used by
Rodrigues et al. (2013) in these other taxa but certainly should prove useful.
Thus, the authors only “proof” for differences are the “representative”
spectrograms included in their Figure 3. I use quotation marks in
representative because we are just forced to believe or to doubt if those
spectrograms are really typical or not (as no data on variation is included in
the article). In fact, spectrograms provided for fuscicapillus and layardi
are clearly different but, as discussed for Boesman (2016), such differences
depend on how they are presented. As Boesman, I tracked both recordings in
Macauley Library and they clearly are not as different as presented in
Rodrigues et al. (2013) (Figure 2).
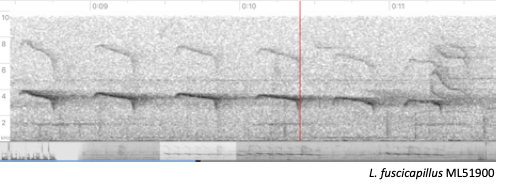
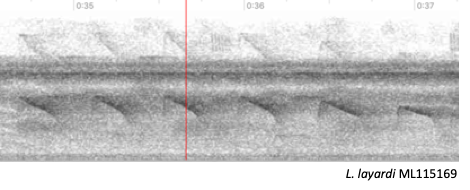
Figure 2.
Spectrograms of songs of L. fuscicapillus
(left) and L. layardi (right).
“Song by fuscicapillus shown in Figure 2
(ML51900) is the same depicted in Figure 3 of Rodrigues et al. (2013). It is
not just different but it is also more representative of this taxon songs
(which could be checked reviewing recordings available both in Macaulay Library
and XenoCanto). On the other hand, I am showing a recording of layardi that is not the same as shown in
Rodrigues et al. (2013) as the one shown by the authors is from a bird on
breeding condition (courtship, display or copulation) based on information
provided by the recordist (Curtis Marantz), and such condition might exaggerate
some of the note shapes. When both song spectrograms are visually compared it
is clear they are not as different as portrayed in Rodrigues et al. (2013).
“Consequently,
based on morphological and vocal information, Rodrigues et al. (2013) do not
provide enough information to make any evaluation of the status of the complete
L. albolineatus complex and, more
specifically for this proposal, of the separation of fuscicapillus and layardi.
Boesman (2016) and Minns (2016) qualitative comments are not complete either.
Besides statements/descriptions suggesting that vocalizations of fuscicapillus and layardi are not clearly distinguishable, no formal vocal analysis
was included. Major comparisons are done focusing mostly on albolineatus and fatimalimae, but I am missing an analysis among all taxa, and
quantitative and qualitative comparisons among duidae, fuscicapillus and
layardi are needed. Such comparisons
should have a clear criteria to define what we consider a species under the BSC
used by this committee. Vocalization thresholds are not fixed and could vary
depending on the species groups compared. Additionally, some taxa might not
differ in their loudsongs but their calls might be important to keep
reproductive isolation.
“The last point I would like to
mention refer to the phylogenetic hypothesis and its interpretation. In this
proposal, Boesman states “it appears that fuscicapillus
and layardi are the two (sister)
taxa which have the smallest genetic divergence”. Similarly, Rodrigues
et al. (2013) indicated in their Figure 1 “numbers refer to posterior
probabilities values and genetic distances between sister groups
associated with the labeled nodes”. It is important to understand that the
phylogenetic hypothesis included in Rodrigues et al. (2013) do not show
any sister relationships among fatimalimae,
duidae, fuscicapillus and layardi.
It only shows albolineatus as
potential sister taxon to a unresolved group (polytomy) formed by these four
taxa. Additionally, regardless of the relevance (or lack of) of genetic
divergence for taxonomic decisions, fuscicapillus
and layardi do not show the smallest
genetic divergence, as such divergence is the same among all taxa included in
the polytomy, as clearly indicated in Figure 1 of Rodrigues et al. (2013). The
lack of evidence for sister taxon relationship between fuscicapillus and layardi
is a clear limitation to lump these taxa into a single species. What if fuscicapillus, for example, turned out
to be closer related to duidae? Would
we be asking the same questions?
“As molecular variation within
lineages was not shown by Rodrigues et al. (2013), I downloaded from GenBank
all sequences used in this study plus the ones used by Arbelaez-Cortes et al.
(2012, Zoologica Scripta) for their study on Lepidocolaptes molecular systematics, to run a Maximum Likelihood
phylogenetic analysis (Figure 3). The phylogenetic hypothesis for the genus Lepidocolaptes, using same gene and
sequences (plus the ones for the rest of species), clearly suggests that the
story is more complex. L. albolineatus
might be related to other taxa within the genus though there is a total lack of
resolution at the base of the tree with few robust relationships among taxa.
Each lineage, however, is strongly supported, and layardi shows a well-supported structure with two groups diverging
in 2% for that gene (ND2). Unfortunately, I don´t have information about
localities to explore if there is any geographical signal in such pattern.
Thus, not only the evolutionary history of this group is more complex than we
thought but also the lack of strong support for any phylogenetic relationships
suggests caution before any taxonomic rearrangement is proposed, both for the L. albolineatus species complex and,
particularly, regarding the taxonomic status of both fuscicapillus and layardi.
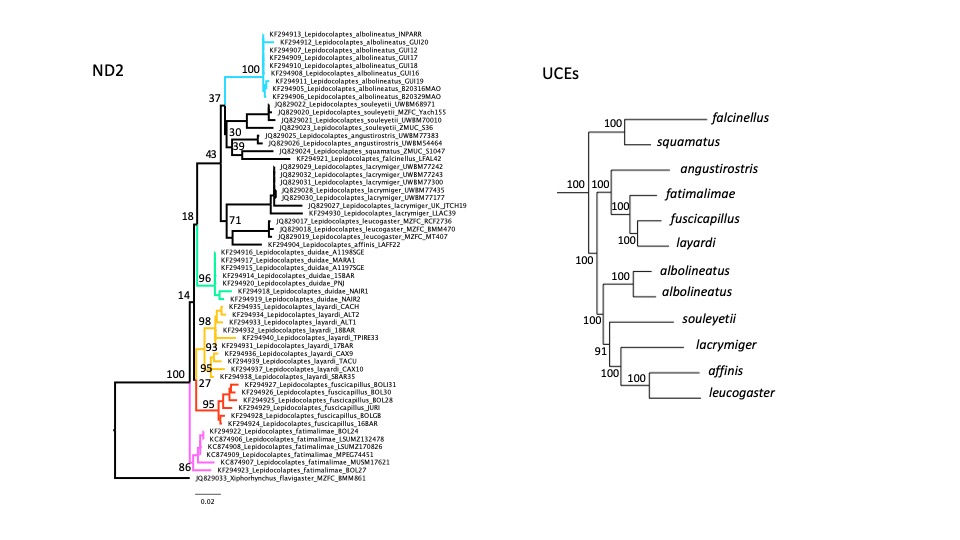
Figure 3. Phylogenetic hypotheses of the genus Lepidocolaptes based on the ND2 gene
(left) and Ultraconserved Elements (UCEs) (Harvey et al. in review; right). The
ND2 hypothesis shows in color each of the five currently recognized taxa for
the L. albolineatus species complex.
Both hypotheses show bootstrap support values based on Maximum Likelihood
analyses.
“Additional but still unpublished
evidence is provided by upcoming suboscine phylogeny by Harvey et al. (in
review) (kindly provided by Mike Harvey). A Maximum Likelihood analysis based
on genomic sequences (UCEs) resulted in a well-resolved phylogeny showing a
polyphyletic L. albolineatus species
complex and suggesting that fuscicapillus
and layardi are sister taxa (Figure
3). This phylogenetic
hypothesis is, by the way, congruent with the previous two-species taxonomy of
this group (albolineatus/fuscicapillus; Cory & Hellmayr 1925,
Catalogue of Birds of the Americas). Though
genomic data are not immune to analytical problems, and this phylogeny only
includes one individual/taxon, at least we have some evidence that fuscicapillus and layardi might be sister taxa. I say “might” because until duidae is not included we are missing an
essential part of the picture.
“In
summary, splitting or lumping fuscicapillus
and layardi into two or just a single
species requires evidence that has not been available to review. A thorough
morphological and vocal analyses should be made available considering the
geographical distribution of each taxa, contact zones (if any), and the potential
geographical variation suggested by the ND2 gene hypothesis (for layardi). Evidence to split the complete
species complex was not thorough or available and, definitely, neither
morphological nor vocal evidence were available to consider these two taxa as
different species. Lumping them back into a single species also requires
evidence. Even when more thorough analyses might eventually show they do not
differ either morphologically or vocally, there is no available information
indicating fuscicapillus and layardi are sister taxa (until duidae is included in phylogenetic
analysis). Additionally, geographical distribution of layardi is congruent with many other Amazonian taxa and a large
genetic divergence (even for just one mitochondrial gene) suggest some sort of
isolation, a pattern that needs to be further explored (looking for potential
causes of the lack of congruence between phenotypic/molecular characters).
Unfortunately, there is no basis to keep the current treatment (i.e., different
species) and lumping them appears to be the available practical solution for
the time being (meaning that a change in taxonomic status is not based on
demonstration they belong to one species or that each lineage does not
represent a single species).”
Comments
solicited from Mario Cohn-Haft: “I don’t really have much of substance to add on this
one based on my personal experience. Basically, I believe that the songs are
virtually indistinguishable between layardi and fuscicapillus. But as I reread the Rodrigues et al. paper and
other evidence called to bear, I am a bit mystified by:
“1)
the rather different looking sonograms between the two in the Rodrigues et al. HBW
paper— I’m curious (but not enough to look them up) to hear those particular
cuts that are graphed. The fuscicapillus
is pictured very faintly even in the strongest parts of the song, so it’s
actually conceivable that the higher peak in layardi notes are actually
made in that cut too, but were filtered out of the figure! And even if not, not sure how different that
would sound. In any case, most cuts of
both spp. sound pretty much identical to me, and in my days of greater activity
in se Amazonia, I never made a distinction between e and w of the Tapajós
birds, but was acutely aware that the 4 quadrants of the Amazon (formed by Negro,
Solimões/Amazon, Madeira) each had different sounding albolineatus types—
i.e., the 4 taxa that everyone’s comfortable with.
“2)
I don’t understand the phrase "the fuscicapillus group is vocally
heterogeneous, with taxa fuscicapillus and layardi being very
distinct vocally, hence suggesting that the polytypic L. albolineatus may
include more than a single species” in the intro of Rodrigues et al. Is that supposed to mean that the two differ
from one another, or that they (as a unit) differ from the rest? It seems like an odd statement to put in the
intro as is, and yet seems to have been picked up in the SACC deliberations
repeatedly as if it were a conclusion.
“It’s
not all that unusual for allospecies to have similar songs, but to differ in
calls or in overall repertoire or in the use of the same repertoire in
different behavioral contexts, so having the same song doesn’t put me off
species distinctiveness automatically. But
it definitely puts up a red flag.
“The
genetic difference, however, seems convincing to me. Yes they’re the most recent split. but the
data, as presented, indicate two nice reciprocally monophyletic groups with a
substantial percent divergence. That
suggests they really are not mixing and that to me is pretty strong evidence of
species status. But similar cases have
proved with subsequent genomic analyses to have lots of gene flow, and
reinterpretation has been necessary. So,
I’d feel a lot more comfortable seeing genomic analyses.
“Sampling
is obviously an important issue. In the
pdf of the article, I can’t make out from the map which localities were
sequenced. I’d sure like to see more
samples and from more strategic localities. Even not being able to distinguish the
different kinds of sample points in the figure, I can see that the potential
contact zone was not well sampled, which seems like an important lacuna for
concluding anything.”
“Finally,
even if the species hold up as distinct, it’s not clear whether the names are
right. The type locality of fuscicapillus,
if I’m reading the map right, is from near to where a contact zone should be
expected. Interestingly, it's also not
far from localities with both (!) fatimalimae and fuscicapillus. That sympatry in itself is interesting (as
mentioned by one of the reviewers of the proposal) and could point either to
sympatry strengthening the species status of the former, or to a zone where potentially
all 3 taxa come into contact and could mix, and where modern analyses should be
focused.
“In
all, rejecting species status for layardi is basically a conservative
move and seems reasonable to me. Not
because there’s strong evidence that it’s not a species, but because there is
still a lack of strong evidence that it is.”
Comments
solicited from Jason Weir: "I have not analyzed the contact zone between Lepidocolaptes [fuscicapillus] layardi and L. [f.] fuscicapillus,
so I can only offer an opinion based on
other contact zones we have analyzed for woodcreepers in this region.
Subspecies of Dendrocincla fuliginosa, Glyphorynchus, as well as Xiphorhynchus
elegans/spixii that come into
contact in this headwater region (Rio Teles Pires) all have very narrow hybrid
zones. I consider them all to be excellent biological species on the basis of
intrinsic postzygotic isolation rather than premating isolation which, based on
vocal divergence is presumably lacking (Dendrocincla
fuliginosa) or weak to moderate
(Xiphorhynchus elegans/spixii), and does not prevent extensive interbreeding
at the contact zone. Each of these species pairs is > 5% GTR-gamma distance
in cytochrome b. In contrast, genomic data I generated for a younger pair
(<4% GTR-gamma distance in cytochrome B) of woodcreeper subspecies from the Xiphorhynchus guttatoides group
demonstrates a very broad hybrid zone here and while they would be considered a
phylogenetic species and have been considered species in the past, it certainly
is not reproductively isolated. Contact
zones are unlikely to occur for all Amazonian taxa and studying those that do
exists takes considerable effort, thus I think it is prudent to extrapolate
evidence we have for the timeline to postzygotic reproductive isolation observed
in currently published hybrid zones to taxa like L. [fuscicapillus] layardi.
Given my hybrid zone data, I suspect
that even in the absence of vocal differentiation, I would recognize
woodcreeper taxa in this region that are > 5% diverged as likely possessing
strong intrinsic postzygotic isolation and I would have no hesitation defining
them as likely representing biological species on this basis. Less than 5%
divergence would require more careful consideration and in the absence of
strong vocal differentiation I would favor taxonomic lumping."
Comments from Pacheco: “YES. I agree that the split of
these two taxa was a misinterpretation and that the available vocal data points
to a very close relationship.”
Comments from Bonaccorso: “YES. The evidence supports
lumping these two taxa into one.”
Comments from Zimmer: “YES, but with reservations about how to
interpret the genetic data and phylogenetic hypothesis, given that, as
correctly pointed out by Jorge, the paper by Rodrigues et al. (2013) does not
resolve the issue of sister relationships among fatimalimae, duidae, fuscicapillus and layardi. The evidence
presented does not unambiguously make the case for splitting fuscicapillus and layardi, but neither does it prove that they should be lumped. We are left in the position of not knowing
what we don’t know. I think the
interpretation that Rodrigues et al. (2013) were focused primarily on
documenting the distinctiveness of fatimalimae
is correct. Remember that there was
confusion in the literature over the subspecific identity of populations from
west of the Madeira – these were attributed by Peters to fuscicapillus, despite the fact that fuscicapillus was described in 1868 from Mato Grosso, east of the
Madeira. The population from the Madeira-Tapajós interfluve, was described in
1919 from Porto Velho, on the east bank of the Madeira in Rondônia, as
subspecies madeirae. Andy Whittaker hit on the fact that birds
from the west bank of the Madeira in Brazil sounded the same as birds on the
rio Javari (Brazil-Peru border) [all purported to be fuscicapillus according to Peters] but distinctly different from
birds from the east bank of the Madeira in Mato Grosso and Rondônia, the
respective type localities for fuscicapillus
and “madeirae”. Songs of birds from those type localities
sounded identical, and it didn’t make sense from the standpoint of biogeography
that there were two identical sounding subspecies occurring on the east bank of
the Madeira in Mato Grosso and Rondônia.
Marantz, et al. (2003 in HBW Volume 8), had apparently reached that
conclusion, and restricted the range of fuscicapillus
to west of the Madeira, seemingly overlooking that the type specimen was from
the east bank. All of this led Whittaker
and his eventual co-authors to conclude that “madeirae” was a junior synonym of fuscicapillus, that it occurred only east of the Madeira, and that
the form with a different voice that was widespread west of the Madeira, lacked
a formal name, and that is the taxon
novum which Rodrigues et al. (2013) described as fatimalimae. I would agree
with Mario Cohn-Haft that the Lepidocolaptes
occurring in the four quadrants of Amazonia:
1) albolineatus in the NE; 2) duidae in the NW; 3) fatimalimae in the SW; and 4) fuscicapillus + layardi in the SE all
sound distinctly different from one another (although there was no adequate
analysis in Rodrigues et al. [2013] to demonstrate this), but that there is no
real distinction between the voices of birds on either side of the Tapajós (fuscicapillus to the west and layardi to the east). Given that, I’m fine with taking a
conservative approach and lumping layardi
into fuscicapillus (which has
priority), despite my aforementioned reservations regarding the genetic data
and associated phylogenetic hypotheses.”
Comments by Lane: “YES. I think Peter has laid out
a strong case for lumping L. layardi into L. fuscicapillus.”