Proposal (906) to South
American Classification Committee
Treat Pachyramphus
[albogriseus] guayaquilensis as a separate species
Effect on SACC: This would raise to species rank a taxon that is endemic to W
Ecuador and NW Peru; it is currently treated as a subspecies of Pachyramphus
albogriseus.
Background: This
taxon was described by J. T. Zimmer (1936, American
Mus. Novitates) as a subspecies of P. albogriseus,
with the following diagnosis:
“Similar to P. a. salvini of
northern Peru' but somewhat smaller; tail of males with more gray, the subexternal pair of rectrices having the basal two-thirds
definitely gray or grayish on both webs. Females not certainly distinguishable
except by smaller size.”
With such a seemingly
minor difference in plumage and size, the taxon went essentially unremarked
after its description. Wading through
Zimmer’s text only leaves me more confused about the true range and characters
of guayaquilensis.
Zimmer described its geographic
distribution as:
“Western
Ecuador in the Tropical Zone surrounding the Gulf of Guayaquil; probably
extreme northern Peri in the same habitat.”
And the specimens Zimmer assigned
to guayaquilensis were from the following localities:
“ECUADOR: Chimbo, 1 male (type); Guayaquil, 1 male; Isla Puna, 2 males; Punta Santa Ana, 1 male; Chongocito, 1 female; Daule, 1 female; San Javier, 1 female”
Ridgely & Greenfield (2001; Ecuador book)
barely mentioned guayaquilensis except to note that they were only
moderately certain that some recent specimens from NW Ecuador were P. a.
salvini or guayaquilensis.
In fact, wading through Zimmer’s original text indicates that he was
somewhat uncertain about specimens from the Andes of NW Ecuador (Mindo in Pichincha).
Schulenberg et al. (2007; Peru book) noted what
appear to be differences in the transcriptions of songs of guayaquilensis
and P. a. salvini but did not comment further (as was their style in
many cases in which species limits were obviously in question, e.g. Cnemotriccus).
However, perhaps it’s just me, but I
really don’t hear much of a difference – below are 3 online recordings from as
close to the type localities as was available.
I would transcribe them all as basically “chu-chu-WEEE?, chu-chu-WEEE?,
chu-chu-WEE?, and I see little qualitative difference among 5 of the 6
sonograms in terms of general syllable shape, pitch, pace.
Sonograms:
guayaquilensis:
https://www.xeno-canto.org/259057 (Guayaquil; John
Moore)
https://www.xeno-canto.org/260979 (Manabí; Mitch Lysinger)
https://www.xeno-canto.org/251294 (Loja; Niels Krabbe)
salvini:
https://www.xeno-canto.org/54569 (Húanuco;
Dan Lane)
https://www.xeno-canto.org/150850 (San Martín; Herman
van Oosten)
https://www.xeno-canto.org/227116 (Cajamarca; Peter
Boesman)
As currently circumscribed, Pachyramphus albogriseus consists of 5
subspecies:
• ornatus (Talamanca-Chiriquí
mountains of Central America)
• coronatus (Santa Martas)
• nominate (Andes of NE Colombia,
Venezuela)
• guayaquilensis (lowlands of Esmeraldas
to Piura)
• salvini (Andes (E Nariño to
Cuzco; type locality = Chusgon [La Libertad, Peru])
What stands out
to me is that guayaquilensis appears to be a lowland taxon, in contrast
to the other four, which are montane. It
also appears possible that they are elevationally parapatric in NW Ecuador,
which really smells like separate species.
Nonetheless, in terms of plumage and voice, I can see why guayaquilensis
does not seem to have attracted any attention in terms of taxon rank.
New information: My attention was drawn to this when perusing the trees in Musher
& Cracraft (2018): They found that P. albogriseus was
not monophyletic. In their mtDNA tree (Fig. 2; note that they used PSC, thus
elevating all subspecies to species rank), their 8 samples of guayaquilensis
were not even in the same clade as the three Pachyramphus albogriseus
subspecies sampled (nominate, salvini, ornatus):
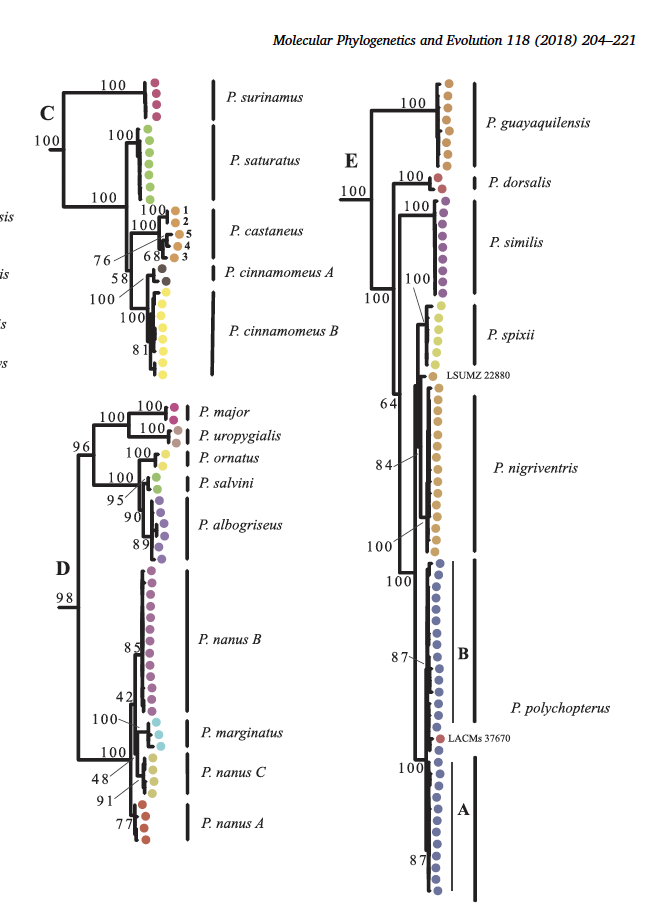
They found the same result in their UCE tree
(Fig. 3):
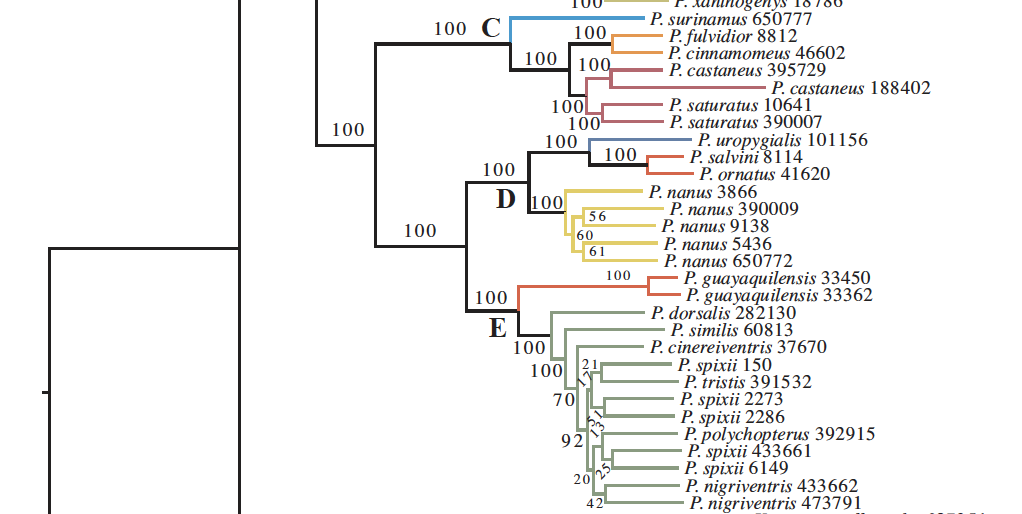
Curious about the localities for the samples
assigned to each taxon, I went to their Supplementary Material online, where
thankfully, following best practices for this kind of research (and in contrast
to many papers), Musher and Cracraft provided the following (rearranged by me
in a more sensible geographic sequence):
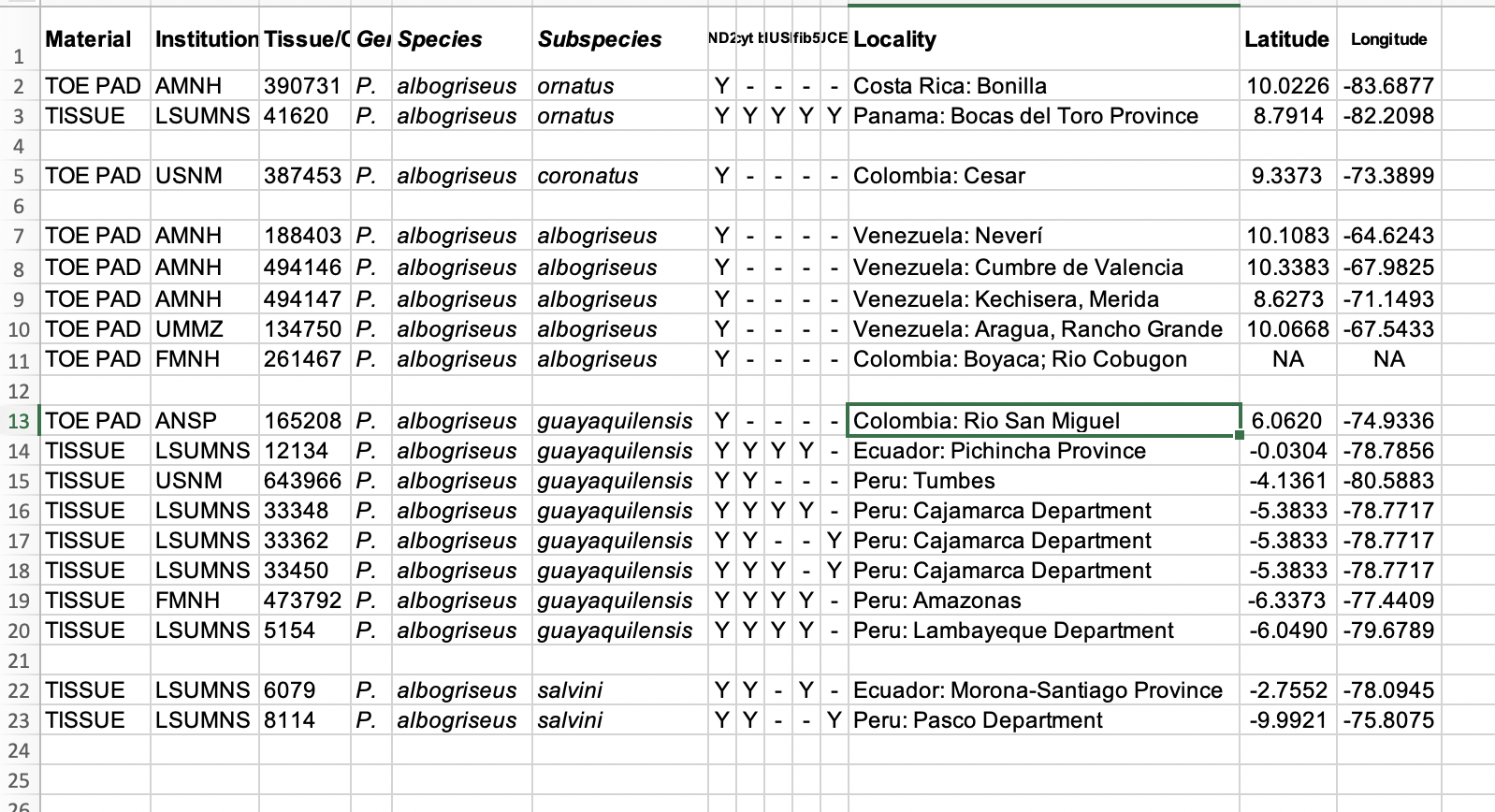
Note that the samples they considered guayaquilensis are not just lowland tropical zone, as
described by Zimmer but include one from the EASTERN slope of the Andes in Nariño,
Colombia – Río San Miguel is a tributary of the Putumayo. Or is it?
The Lat-Long given actually maps to a locality about 70 km ESE of
Medellin in what appears to be Antioquia on the east slope of the Central Andes
(which I assume to be an error). Using
the coordinates provided, the Ecuador sample comes from a locality on the west
slope of the Andes NW of Mindo at ca. 1575 m. Also, the Amazonas, Peru, locality is
interesting – those coordinates map to about 50 km ESE of Chachapoyas, east of
the Marañon at about 2250 m, i.e. what I think might be closer to the
traditional distribution assigned to salvini. Seasonal movements? That these 8 samples all
clustered together tightly in Fig. 2 is reasonable justification for
considering them the same taxon, regardless of traditional distribution
assigned to guayaquilensis. Unfortunately, no toepad samples were used
from specimens from near the type localities of guayaquilensis or salvini.
Puzzled by all
this, I sent the first draft of the proposal to Dan Lane and Tom Schulenberg to
see if I was missing something. Both had
also been puzzled by this finding, and in fact, unbeknownst to me, Tom had
already covered this problem in his “Split, Lumps, Shuffles” column in
Neotropical Birding #26, the text of which I copy here (and made me wish I’d
read this first):
“The real surprise coming out of
this research is something else – something completely unexpected. Musher and
colleagues find that most populations of Black-and-white Becard P.
albogriseus are sister to Grey-collared Becard P. major, but one
group of Black-and-white turns out to be related instead to White-winged Becard
P. polychopterus. This is a pattern that they found with several
different genetic markers, a very strong signal that this result is not an
artefact (i.e., a peculiarity due to an odd pattern of inheritance of one
particular gene), and that this group represents a separate species. This would
be quite surprising, since the subspecies of Black-and-white Becard differ from
one another in only the most minor of details – so if a species, it is a very
cryptic species indeed. Musher and colleagues nonetheless blithely split this
lineage as P. guayaquilensis, which is where the complications begin.
“Their
samples identified as P. guayaquilensis come not only from western
Ecuador, the range generally attributed to that subspecies, but also from sites
as far afield as the east side of the Andes, from southern Colombia to northern
Peru – regions from which ‘true’ Black-and-white Becard also occurs. Some
observers (the late Paul Coopmans, in Ridgely & Greenfield 2001, and Dan
Lane, in Schulenberg et al. 2010) have described two different songs of
Black-and-white Becard. If there is any connection between song types and
genetic groups, however, Musher et al. will not be the ones to tell us:
they do not discuss the issue at all, much less point to any individual becards
for which they have both an audio recording and a genetic profile. They leave
us with a real headscratcher. Perhaps if we’re lucky, some other researchers
will return to this topic, and sort it all out for us.”
In conclusion, I
have no real conclusion. That the guayaquilensis
samples pertain to that taxon, and that they are part of a different clade of Pachyramphus
species, seem undeniable from Musher & Cracraft. But why then would guayaquilensis be
marginally distinguishable from salvini in plumage and voice? None of this makes any sense to me.
Recommendation: I wrote up this
proposal because I thought it was going to be straightforward and because I
wanted to study this case, which was completely unfamiliar to me. Well … I was wrong, and it likely would have
been better for someone more familiar with the specifics. I have no recommendation, and will vote after
seeing what others say.
Note on English names: I’m waiting to see
whether the proposal has a chance of passing before I put any work into
this. Without any diagnostic plumage or
vocal characters, and with a geographic range that would extend far beyond the
original concept of guayaquilensis being restricted to lowlands of W
Ecuador and NW Peru, I see no obvious choices at the moment other than an
unsatisfactory Guayaquil Becard.
Van Remsen, February
2021
Comments
from Stiles:
“NO. Given the uncertainties regarding localities,
song differences or not, minor plumage differences and the lack of association
of specific specimens for which both song and genetic data were reported, I
must agree that separating of P. guayaquilensis is definitely
"unsatisfactory" on present evidence, so NO, for now.”
Comments
from Lane:
“NO. Despite having discussed this situation with
various folks (including Luke Musher, Tom Schulenberg, and Van Remsen), I have
also ended with my head spinning with the conflicting information of
phylogenetics, voice, and plumage as Van has outlined above. In response to an
initial query by Musher as he was about to publish in 2016, I told him the
following:
"Unfortunately, I have rather limited experience with P.
albogriseus, and have only had a couple of opportunities to record its
voice. This sounds like the sort of thing that Paul Coopmans would perhaps have
figured out, so if it is not touched on in Birds of Ecuador (where many of Coopman's
observations were published), you're probably on novel ground! The sample size
I was working with when writing the voice accounts for Birds of Peru was small,
and I'm not sure that I represented the differences between the two taxa well.
Your comments about the distinctiveness of the east and west slope populations
are revelations to me, and I cannot offer anything new to you to support or
deny the patterns you have uncovered. Checking the LSU museum holdings to see
if they are in line with the differences you mention, I see that we have,
overwhelmingly, guayaquilensis (several identified as "salvini"
as you say) with only two definite salvini from Pasco and Ayacucho [Comment
inserted Feb 2021: Musher had outlined that he'd noticed strong size
differences, particularly obvious in bill size, between what he was calling guayaquilensis
and salvini, with an additional character being the female crown pattern
as I mention in the next sentence]. The female from Ayacucho has a distinct
black band outlining the rear of the crown behind the eyes, the crown is warmer
chocolate (darker, colder brown in guayaquilensis), the cheeks are not
as gray as guayaquilensis females, and the wings have far brighter,
warmer cinnamon edgings. Our guayaquilensis specimens are from Tumbes,
Piura, Lambayeque (all west slope), but also from the Chinchipe and Utcubamba valleys, as well as La Libertad, which are
"east slope" although they drain into the Maranon River, which often
captures "west slope" taxa."
So, honestly, this is still where things stand for me. As Van
indicates with his search of vocalizations, it is unclear if the two groups
differ in voice... but given few, if any, recordings are linked to specimens,
it could be that there are two overlapping taxa in the range usually assigned
to "salvini" and the vocalizations being associated with that
taxon may not actually correctly named. Clearly, something weird is happening
here, and it may require a student dedicated to this project doing some
fieldwork targeting the complex to clarify what it is. Sadly, at least in my
experience, P. albogriseus is far from easy to find, much less record,
so it will be an uphill battle for whomever tries to take it on! One thing that
will be necessary will be to verify that the type specimens of guayaquilensis
and salvini each truly fall in the clade assigned their respective
names in the tree above! Until such a project is able to make better sense of
things, I will have to vote NO.”
Comments from Areta: “YES. The genetic data strongly show that we are in
the presence of two different species-level taxa. I examined the recordings
looking for vocal types, and what I found is that there appear to be two main
song types. A rapid, explosive type and a slow, mellow type, which match (to my
understanding) the descriptions assigned by Schulenberg et al. for guayaquilensis (rapid) and salvini (slow).
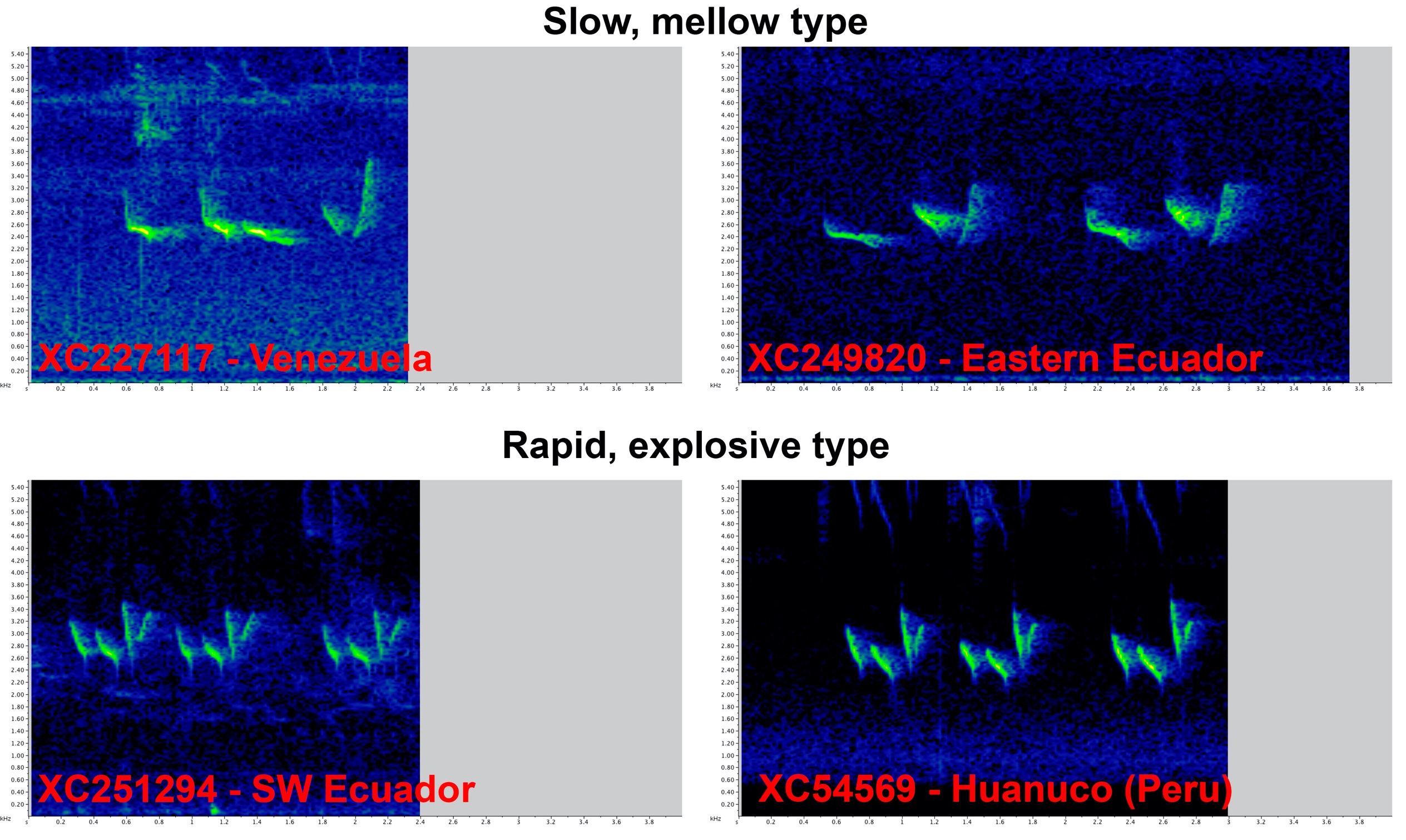
“According
to this assessment, the voices that Van shared in his proposal would be:
Rapid,
explosive:
https://www.xeno-canto.org/259057 (Guayaquil; John
Moore)
https://www.xeno-canto.org/260979 (Manabí; Mitch Lysinger)
https://www.xeno-canto.org/251294 (Loja; Niels Krabbe)
https://www.xeno-canto.org/54569 (Huánuco; Dan Lane)
https://www.xeno-canto.org/227116 (Cajamarca; Peter
Boesman)
Slow, mellow:
https://www.xeno-canto.org/150850 (San Martín; Herman
van Oosten)
“When
plotting these mentally on a map, the slow, mellow type is the only one to
occur in the area of nominate albogriseus (e.g., all Venezuela
recordings), while the rapid, explosive type is the only one known to occur
near the type locality of guayaquilensis, but note also that this vocal
type may be the one occurring at the type locality of salvini and both
vocal types can occur fairly close to each other in Ecuador and Peru, although
apparently the slow type is always to the east of the rapid one.
“So,
to me, there are two species here, one that can be safely called albogriseus
based on genetics and vocalizations (slow, mellow type), and a second one that
could be called either guayaquilensis or salvini depending on how
true salvini vocalizes (it seems certain that guayaquilensis
gives the rapid, explosive type). I am not sure that the specimens assigned to salvini
by Musher & Cracraft 2018 actually pertain to salvini (type locality
Chusgon, Peru at -8.108206 -78.070969), and this is
the poignant point that need to be sorted to decide which name must be applied
to the rapid, explosive type: guayaquilensis or salvini?. The
fact that Zimmer described guayaquilensis
as very similar to salvini, makes me
suspect that they both are rapid singers.
“Intriguingly,
the coordinates from the "salvini" specimen from Peru (-9.9921, -75.8075 plot to Huánuco not Pasco
as stated in the supplement), very close to where XC54569 (rapid) was recorded.
While the other "salvini" specimen from Ecuador (-2.7752,
-78.0945) comes from an area in which only the slow type song (e.g., https://macaulaylibrary.org/asset/245718) is known. What this
suggests to me is that the "salvini" samples in Musher &
Cracraft pertain indeed to the slow, mellow type (consistent with their
phylogenetic placement) but this does not necessarily mean that true salvini
pertain to this vocal type. Indeed, based on where Chusgon
is placed, and based on the fact that slow vocalizations seem to occur on the
eastern slope of the Andes in the southern part of its distribution while birds
from Piura and Cajamarca deliver the rapid type, it seems to me that true salvini
might pertain to the rapid type, thereby having priority over guayaquilensis
as the species name for the fast singers. It seems that "the" problem
here is which name to apply to the rapid type. If this assessment is correct,
then what we would have is:
Pachyramphus
salvini
(including guayaquilensis, either as
the same ssp. or a different one): note that this would demand that all the
slow singers traditionally assigned to salvini,
be placed together with albogriseus.
Pachyramphus
albogriseus
(including the rest of the subspecies)
“The
situation with the morphological distinctions needs more assessment, and critical
comparisons of the type specimens (notably of salvini and guayaquilensis
to albogriseus, and to sequenced
specimens) are needed. However, because birds from both genetic groups are
housed at LSUMNS, we can benefit from examining pictures and testing these
ideas.
“I
spent a full morning examining this, but I am sure that people with more field
experience and specimens at hand can provide arguments to refute or corroborate
the ideas here proposed.
Comments
from Bonaccorso:
“NO. I think this one requires a better
understanding of what is what, how it sings, and where it lives (not only in
terms of region but also habitat). I think this is an excellent opportunity for
a student to do a thorough study, including field, specimen, and acoustic components.”
Comments
from Stiles:
“A real conundrum.. As Nacho noted, the genetic data indicating two species
seem irrefutable, but morphological, geographic and vocal differences are to
say the least, confusing and incomplete, and if guayaquilensis is indeed
separable, we don’t know what its Latin name should be: if found to include salvini,
the name guayaquilensis becomes a
synonym. So, I will continue to vote NO for now, in hopes that we might soon
have a few cases of specimens with both voice recordings and tissue
samples from well georeferenced critical localities to unscramble the knots.”
Comments from Claramunt: “YES. Genetic data strongly suggest there are two species involved, and Musher & Cracraft also describe the plumage traits that differentiate them. They may also differ vocally, as Nacho suggested. There may be some uncertainty now regarding the precise geographic limits of the two species and even the allocation of the described subspecies to each of the species, but I think that the fact that there are two species involved is solid.”
Comments
from Robbins:
“Unexpected, fascinating, and certainly mystifying. Clearly, there is at least an additional
species currently considered as albogriseus. As pointed out, to get at
what name should be applied to this species is highly debatable and appears
that the holotypes of at least salvini and guayaquilensis are
going to need to be sequenced and compared to unequivocal genetic material of albogriseus.
So, until things are clarified, probably best to put this on hold; thus, I vote
NO for now.”
Comments
from Pacheco:
“NO. Molecular data suggest that there are two
species involved. However, there are doubts about the affiliation of some of
the samples. The relationship between guayaquilensis and salvini remains to be
resolved (as Gary pointed out), which interferes with the split.”
Comments
from Remsen:
“NO. Clearly, something is amiss here,
and this needs to be sorted out with additional work on the genetic samples,
type localities, and vocalizations. Obviously,
two species are involved here, but the geographic limits need to be worked out
and other issues resolved before changing the classification.”