Proposal (908) to South
American Classification Committee
Split
Barn Owl (Tyto alba) into three species
Note from Remsen: Below is a proposal that was rejected by NACC and is posted here
with permission from its authors. If it
passes, it would change the classification of our species to: American Barn-Owl
(Tyto furcata).
Background:
Taxonomic references currently recognize from
one to four species in the Barn Owl (Tyto
alba) species complex. The AOS recognizes one cosmopolitan species (AOU
1998), as does HBW, which additionally groups 28 subspecies into eight species
groups (del Hoyo & Collar 2014). Clements et al. (2016) differentiate T. deroepstorffi (Andaman
Islands) from T. alba, whereas the
Howard and Moore Checklist recognizes three species: T. alba, T. deliculata, and T.
deroepstorffi (Dickinson & Remsen Jr. 2013).
The IOC (Gill & Donsker 2018) recognizes four species of barn owl:
· T. alba (Western Barn Owl), 10 subspecies, widespread in Africa and Europe
· T. furcata (American Barn Owl), 12 subspecies, widespread in North, Middle, and
South America
· T. javanica (Eastern Barn Owl), 7 subspecies, distributed from south and
southeast Asia to Australasia and southwestern Pacific
· T. deroepstorffi (Andaman Masked Owl), from the Andaman Islands
A different subset of three allopatric species
has been recognized based on molecular phylogenies derived from sequences of
the mitochondrial cytb gene and the
nuclear RAG-1 gene (Wink et al.
2009): the Common Barn Owl, T.alba, which has ten subspecies distributed in Africa,
Eurasia, and South-east Asia; the American Barn Owl, T.furcata, with at least five
subspecies from North, Central, and South America; and the Australian Barn Owl,
T. deliculata,
with at least four subspecies restricted to the easternmost part of Southeast
Asia, Australia, New Zealand, and Polynesia (Alibadian
et al. 2016). Morphological traits such as overall size, plumage coloration and
pattern, amount of feathering on the tarsus, and power of tarsus and toes have
been proposed to correlate with the three-species subdivision (Alibadian et al. 2016).
New Information:
Alibadian et al. (2016) published a molecular study of systematic relationships
within the Barn Owl species complex, and estimated the timing of divergence
events. Alibadian et al. (2016) analyzed 40 samples
belonging to ten taxa, which included populations distributed across the world
(Table 1). They obtained sequences of three mitochondrial genes, cyt b (620
bp), CO1 (660 bp), 16S (568 bp), and one nuclear gene, RAG-1 (990 bp). They
inferred a maximum likelihood phylogenetic tree (RAxML v. 7.0.4) and a Bayesian
inference tree (MrBayes v. 3.2). They also conducted a molecular dating
analysis (BEAST v. 1.8), an estimation analysis of the ancestral distribution
of the three groups (LAGRANGE), and statistical analyses of ecological niche
overlap (MAXENT v. 2.0, ENMTOOLS).
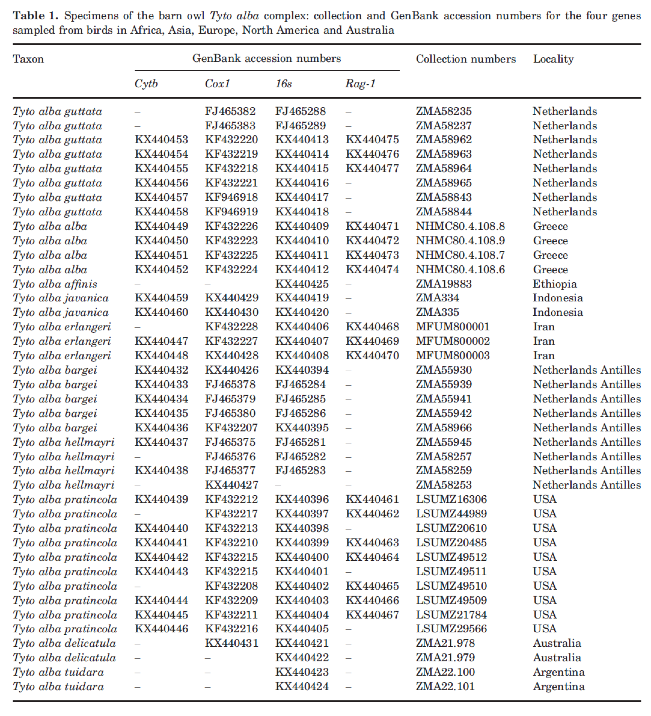
Phylogenetic analyses revealed three main
clades with strong geographic structure. The first group, the furcata clade, included subspecies from
mainland North and South America and Curaçao (T. a. pratincola, T. a. hellmayri, T. a. tuidara,
T. a. bargei). The second group, the alba clade, included subspecies from the
Netherlands, Greece, Iran, and Ethiopia (T.
a. guttata, T. a. alba, T. a. erlangeri, T. a.
affinis). The third group, the javanica
clade, contained samples from Indonesia, India, and Australia (T. a. javanica, T. a. stertens,
T. a. deliculata) (see tree below). The dating
analysis indicated that the Barn Owl complex originated during the Middle
Miocene, and the biogeographical reconstruction suggested an origin in the Old
World. A low amount of ecological niche overlap was estimated among all three
lineages.
Alibadian et al. (2016) proposed that the taxonomy of Tyto alba be redefined and that at least three species should be
recognized. However, because not all T.
alba subspecies were included in the study, the authors established the
species limits based mainly on the geographic distribution of the subspecies
sampled. The authors proposed restricting the specific epithet alba to populations from the
Afrotropical and Palearctic regions to at least eastern Iran. They suggested
elevating the furcata clade
(populations from Nearctic and Neotropical regions, including at least part of
the Caribbean) to species status under the name Tyto furcata. The javanica clade,
including populations from Indonesia, India, and Australia (T. a. javanica, T. a. stertens,
T. a. deliculata), was proposed for elevation
under the name Tyto javanica, because
the name javanica has priority over deliculata.
Uva et al. (2018) published another molecular study of the systematic
relationships of the barn owls and relatives, estimated divergence times using
fossil calibrations, and reconstructed ancestral ranges. Uva
et al. (2018) analyzed 179 genetically different individuals belonging to 16
species of Tyto and 1 species of Phodilus, which
included over 30 subspecies distributed worldwide (see map below). They
obtained sequences of five mitochondrial markers (ND6, CO1, control region,
cytochrome b, and 16S) and two nuclear markers (C-MOS and RAG-1). They inferred
maximum likelihood phylogenetic (RAxML) and Bayesian inference trees (BEAST v.
1.8.4) and constructed a haplotype network of the Common Barn Owl based on the
cyt-b sequences (TCS, POPart).
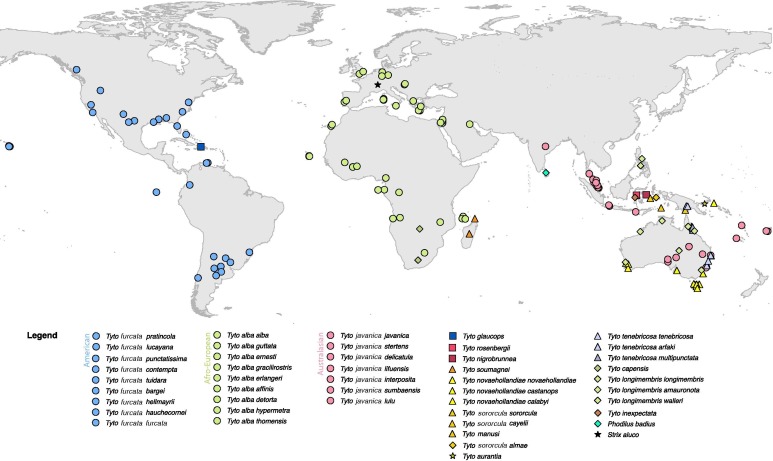
Figure 1. Sample location map - sample
locations, legend following the classification of Gill and
Donsker (2018). When samples were missing precise locations, approximate
coordinates were given.
Phylogenetic and haplotype network analyses
(Figure 3) recovered three main lineages within the Common Barn Owl group,
which differed by 5.82 to 9.33% in cyt-b sequence, supporting the results from Alibadian et al. (2016). The scientific and common names used
by Uva et al. (2018) followed the IOC World Bird
List.
· Australasian clade: samples of the Eastern Barn Owl, T. javanica, from Australia and
Indonesia, including the Sulawesi Masked Owl, T. rosenbergii, T. javanica from India, Malaysia and Java, and T. nigrobrunnea.
· American clade: samples of the American Barn Owl T. furcata, and the Ashy-faced Owl, T. glaucops. This group also includes a subclade of two island
endemics from Galapagos (T. f.
punctatissima) and Hispaniola (T. g.
glaucops).
· Afro-European clade: samples of the crown group of all Western Barn
Owls, T. alba, and the São Tomé Barn
Owl, T. a. thomensis.
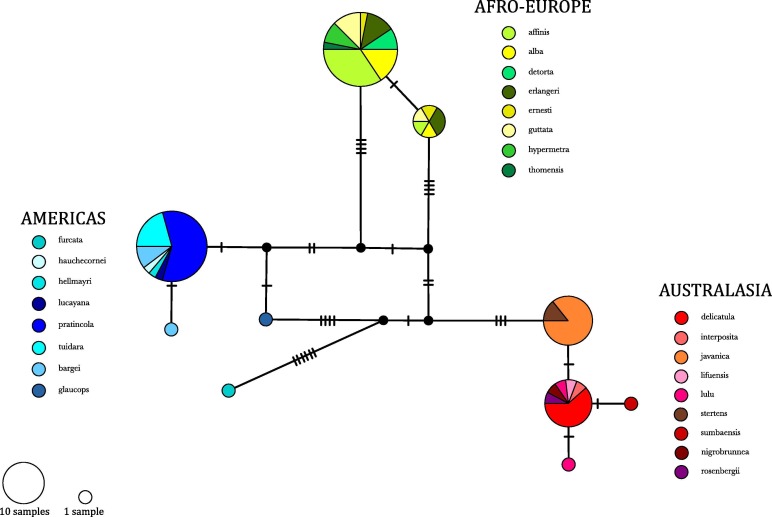
Figure 3. Genetic structure within
the Common Barn Owl group, including all taxa nested within the three major
clades - TSC haplotype networks drawn for cyt-b sequences, following the
classification in Gill and Donsker (2018).
The dating analysis indicated that the Barn
Owl complex originated during the Late Miocene (ca. 6 mya). The biogeographical
reconstruction suggested an origin in the Australasian and African regions.
Uva et al. (2018) concluded that the Common Barn Owl consists of three
evolutionary units, as in Alibadian et al. (2016),
and indicated that three species should be recognized:
· African and European populations: Tyto
alba, Western Barn Owl
· North, Cental and South American
populations: Tyto furcata, American
Barn Owl
· South and southeastern Asian and Australian populations: Tyto javanica, Eastern Barn Owl
Recommendation:
We recommend splitting Tyto
alba into three species to better reflect the evolutionary trajectory of
the clade. The phylogenetic evidence suggests geographic and genetic isolation
of the three lineages (Wink et al. 2009, Nijman & Alibadian
2013, Alibadian et al. 2016, Uva
et al. 2018), in addition to their correlated morphological traits (Alibadian et al. 2016).
(1)
Afrotropical and Palaearctic populations: Tyto
alba, Western Barn Owl
(2) American
populations: Tyto furcata, American
Barn Owl
(3) Eastern
Asian and Australian populations: Tyto
javanica, Eastern Barn Owl
The recommended English names are based on
those proposed by Uva et al. (2018), which are
currently used by the IOC World Bird List (Gill & Donsker 2018).
Literature Cited:
Alibadian M., N. Alaei-Kakhki, O. Mirshamsi,
V. Nijman, and A. Roulin. 2016. Phylogeny,
biogeography, and diversification of barn owls (Aves: Strigiformes). Biological Journal of the Linnean Society
119, 904-918.
American Ornithologists' Union. 1998. Check-list
of North American birds. 7th edition. Washington, D.C.: American
Ornithologists' Union.
Clements J. F., T. S. Schulenberg,
M. J. Iliff, D. Roberson, T. A. Fredericks, B. L. Sullivan, and C. L. Wood.
2017. The eBird/Clements checklist of birds of the world: v2016. Downloaded
from http://www.birds.cornell.edu/clementschecklist/download/
del Hoyo J., and N.J. Collar.
2014. HBW and BirdLife International Illustrated Checklist of the Birds of the
World, Volume 1 Lynx Edicions in association with BirdLife International,
Barcelona, Spain and Cambridge, UK.
Dickinson E.C., and J. V. Remsen
Jr. 2013. The Howard and Moore Complete Checklist of the Birds of the World –
Volume 1 Non-Passerines. Aves Press, Eastbourne, UK.
Gill F., and D. Donsker. 2018. IOC
World Bird List (v 8.1), DOI: 10.14344/IOC.ML.8.1.
Nijman V., and M. Alibadian. 2013. DNA barcoding as a tool for elucidating
species delimitation in wide-ranging species as illustrated by owls (Tytonidae
and Strigidae). Zoological Science 30,
1005-1009.
Uva V., M. Päckert, A. Cibois, L. Fumagalli,
and A. Roulin. 2018. Comprehensive molecular
phylogeny of barn owls and relatives (Family: Tytonidae), and their six major
Pleistocene radiations. Molecular
Phylogenetics and Evolution. https://doi.org/10.1016/j.ympev.2018.03.013
Wink M., A. A. Elsayed,
H. Sauer-Gurth, and J. Gonzalez. 2009. Molecular
phylogeny of owls (Strigiformes) inferred from DNA sequences of the
mitochondrial cytochrome b and the nuclear RAG-1 gene. Ardea 97, 581-591.
Submitted
by: Rosa Alicia
Jiménez and Carla Cicero, Museum of Vertebrate Zoology (to SACC in February
2021)
Comments from Remsen
[this is what I wrote on the NACC proposal]: “--- NO!!! It’s great to see this
proposal because we really need to evaluate all this given the new data, and
IOC and HBW splits. However, this split
is based on degree of genetic divergence of (presumably) neutral loci. It is homologous to bar-coding rationale. Because genetic divergence is measured on a
continuous scale, there is no conceptual basis for calling a split “deep” or
“shallow” or whatever. Yes, there are
three lineages, not surprising for such a widespread species, but are there any
consequences relevant to BSC species limits in the divergence at neutral
loci? Otherwise, all we would be doing
is instituting subjective thresholds on genetic differentiation of loci that by
definition are assumed to be irrelevant to the biology of the birds
involved. The morphological traits said
to be associated with each lineage are interesting but as far as I can tell
just geographic variation largely irrelevant to species limits.
“I cannot think of a single
modern analysis of species limits in any owl (or caprimulgid) that isn’t
anchored in vocal differences. Barriers
to free gene flow are exceptionally strongly correlated with vocal differences
in nocturnal birds. It is widely known
that plumage differences are basically irrelevant in these nocturnal birds (for
obvious reasons, including that some species have strikingly different color morphs). So, how can a proposal in 2018 NOT
concentrate on voice? Until vocal
differences, if any, are elucidated, we shouldn’t even be considering this
split in my opinion, for the reputation of our Committee.
“Not one YES voter has produced a coherent rationale for this split;
some have written that the in press paper (with first and last names of authors
reversed, and at least two run-on sentences … typical MPE) that tipped
the scales, but did not explain why.
Vera et al. presented great data on relationships among Tyto,
with much broader taxon-sampling than the previous paper. However, their taxonomic conclusions are
weakly supported and probably would not have survived a bird journal
review. It’s basically a set of gene
trees, combined, and as stated clearly by the authors, the paraphyly is driven
by mtDNA: “our tree
topology is largely dominated by the signal of mitochondrial markers”. The authors themselves noted: “All these pitfalls have to be kept in mind
for any taxonomic conclusions drawn from newly established phylogenetic
hypotheses in the following, because gene-tree topologies depend on the
combined effects of introgression, incomplete lineage sorting and faulty
taxonomy.” Further, the paraphyly itself is the consequence of
peripheral speciation: three insular taxa currently ranked as species (our
T. glaucops, T. nigrobrunnea from the Sula Islands, T. rosenbergii from Sulawesi). If these insular spinoff taxa did not exist, Then
There Would Be No Paraphyly. For
this and other reasons, use of the monophyly criterion at the population level
is a misapplication of Hennigian principles; Hennig
himself did not use the term monophyly at the species level because, using an
early schematic diagram of incomplete lineage sorting, he showed why species
are not necessarily monophyletic.
“The most interesting finding is that the distinctive T. a.
punctatissima from the Galapagos is the likely sister of Hispaniolan T.
glaucops (yet another Galapagos-Gr. Antilles connection), and in my
opinion, requires a proposal to split punctatissima. Otherwise, all we have is three lineages
corresponding to three major areas.
“The authors’ rationale for the split of Barn Owl into three species is
based (in addition to the paraphyly that they themselves were cautious about)
on the existence of three monophyletic lineages corresponding to three major
regions. This is a typical pattern that
provides no basis for assigning taxon rank.
Although I can’t seem to find where in the text the authors present the
actual genetic distances among them (presented in an unavailable table in
online supplementary material), it is clear that the reasoning is basically
bar-coder rationale. For example, they
regard the split of Osprey into four species as a given because they are
genetically “well-differentiated species”, when the actual % sequence
divergences (1.5-2.6% in Monti et al. 2015) are small; these would be at the
low end of % sequence divergences among populations of Andean or Amazonian
birds separated by rivers.
“The plumage differences among the populations are minor, and perhaps
less than those among subspecies within the three major groups. Regardless, plumages in owls are essentially
meaningless in terms of species limits.
Owls are notorious for color phases, geographic variation, and
individual variation that are not considered relevant to species limits. I think there might be more individual
variation in Glaucidium brasilianum than among these Barn Owl
populations.
“What about voice? A cruise
through what I can find on xeno-canto indicates that the eerie hissing screech
that we are all familiar with in our Barn Owl is also the primary vocalization
of European birds (“Western Barn-Owl”):
England: https://www.xeno-canto.org/186611
Belgium: https://www.xeno-canto.org/235525
Netherlands: https://www.xeno-canto.org/384166
“African birds (also “Western Barn-Owl”) sound similar:
Senegal: https://www.xeno-canto.org/237647
“So do Indian birds (“Eastern Barn-Owl”), at least superficially:
India: https://www.xeno-canto.org/species/Tyto-alba?pg=3
“And just so you know, here’s a
recording from distant Argentina, which sounds to me just like those here in
the USA: https://www.xeno-canto.org/species/Tyto-alba?pg=3
“Perhaps an actual formal analysis would find some consistent,
diagnostic differences among these complex screeches, but the burden-of-proof
in my opinion is showing the differences.
Then, do the differences make a difference to the owls themselves? At the other potential extreme, maybe all Tyto sound alike. If that’s the case, then this needs to be
addressed explicitly.
“Finally, if this proposal somehow passes (despite
not a single committee member explaining why it should), a separate proposal on
English names should be required: is “American Barn-Owl” really the way we want
to go for a species found throughout the Western Hemisphere? Sure, Europeans regard all that as “the
Americas”, but it makes me queasy.
Eastern and Western are out of our purview, but are to be lackluster in
the extreme.
Comments
from Lane:
“NO. I think Van makes some very important points in
his review of the situation here. Overall, I find such genetically driven cases
unconvincing for splits without further correlating characters to back them up.
Voice, as Van points out, seems an obvious character set for a nocturnal group
of birds. That said, I must admit that I am unable to detect an obvious
distinction in the voices of T. "alba" and T. glaucops
(but the latter has only 2 available recordings--of the same individuals--on
XC), two populations we *know* must be acting as good species, given that they
are sympatric on Hispaniola! Of course, Tyto owls seem to have
relatively few useful characters to score in their voices (listening to various
other Tyto on Xenocanto reveals Van's fear:
they pretty much all sound similar), and even being certain of using homologous
vocalizations is difficult (to me, at least). In light of this relative lack of
support of other characters to back up the split up of Tyto alba,
I vote NO.”
Comments
from Areta:
“YES. The genetic evidence shows fairly deep breaks (even deeper than those
between other recognized species of Tyto)
with overall good geographic coverage. As for the voices, I will echo here what
a NACC member wrote in voting on this proposal. Magnus & The Sound Approach
(2015, p. 17; if anyone wants to read it, I have the book) state the following:
‘By contrast, there are
dramatic differences between Common and American Barn Owls. American has much
shorter perennial screeches, typically less than a second long, and in a wide
range of recordings, I have never heard American giving anything remotely like
a courtship screech. Gerrit Vyn is the author of an
excellent CD on North American Owls (2006). When I sent him an example of Common
Barn Owl courtship screeches, he confirmed knew nothing similar from American.
This not only supports separating the two species but also the two kinds of
screeches.’
“At the same time, American Barn Owl has a prominent flight call that is
completely absent in Common Barn Owl. It was Gerrit who recorded the metallic
clicking sound in CD1-06,
which he calls the 'kleak-kleak’' call (Vyn
2006). Unpaired males use it most often (Gerrit Vyn
pers comm), so it must have an important role in mate attraction. Marti et al
(2005) reported that males kleak
in the vicinity of the nest, soon after leaving the daytime roost,
and when approaching with food deliveries. Several other Tyto have similar calls (e.g.,
African Grass Owl T. capensis,
Eastern Grass Owl T. longimembris and Australian Masked Owl T. novaehollandiae). So
rather than being an American invention it seems that Common stopped using this
call and replaced it with courtship screeching."
“The
"kleak-kleak"
is also a common vocalization of birds across Argentina, and it seems
impossible that such a prominent vocalization would have gone undetected within
the range of Common Barn Owl (T. alba
sensu stricto); The Sound Approach would have detected it if it indeed
occurred.
“This
marked vocal differentiation, coupled to genetic data, places the burden of
proof in those willing to maintain the status quo.
“As
for the common English name, American Barn Owl looks good to me. I am American
because I was born and raised in Argentina, much as Tyto furcata tuidara are American.
“The situation with T. glaucops and punctatissima is potentially more complex, and more evidence would
be necessary to make the split (or lump?). However, the proposal does not deal
with this case. This is what Uva et al. (2018) said
about this, which is conservative and reasonable:
"In the American Barn Owl, T. furcata,
we found no differentiation between North, Central and South America.
Phylogeographic structure was only detected between a clade of two insular
taxa, T. g. glaucops from Hispaniola and T. f.
punctatissima from the Galapagos Islands, which were sister to the
remaining continental and insular American Barn Owls. The extreme disjunct
distribution of these sister taxa is even more surprising than those of masked
owl endemics from islands east and west of New Guinea, and given the poor node
support (0.77 PP/66 BS) putative genetic distinctiveness of Caribbean and
Pacific populations needs further confirmation from future phylogeographic
studies. For the time being, a species-level split of T. furcata
from T. alba and T. javanica as previously
advocated by some authors (Aliabadian et al., 2016;
Gill and Donsker, 2018) seems justified, keeping in mind that phylogenetic
relationships of T. f. punctatissima and T. glaucops
are still unclear. Genetic information on the dark island forms from Puerto
Rico (T. glaucops cavatica) and
from the Lesser Antilles (T. g. nigrescens, T. g.
insularis) missing to date are needed for a concise species-level
classification of the American Barn Owl, T. furcata (sensu Gill
and Donsker, 2018) and the Ashy-faced Owl, T. glaucops."
“I want to note that
the problem of what to do with punctatissima
will remain open regardless of whether we split furcata or not. So, I
favor recognizing Tyto furcata as a
separate species, it seems difficult to withhold from this given the vocal and
genetic differences with Tyto alba.
“Additional references:
Robb, M. & The
Sound Approach. 2015. Undiscovered owls, A Sound Approach guide. The Sound Approach.”
Comments
from Bonaccorso:
“YES. After reading Nacho´s comment, I am more
convinced, but we have to admit that if there are voice differences between
these three lineages, the evidence is anecdotal. I agree that neutral-marker
differences (which are pretty high in this case) are not enough to infer
reproductive isolation. However, in a bird with apparently such good dispersal
abilities (think about the islands they have reached), a few dispersal events
between lineages in the last few thousand years would be enough to show
admixture, don´t you think?”
Comments
from Stiles:
“YES to splitting furcata from asio (and javanica) as the
American Barn Owl. The vocal data (especially in a genus like Tyto not
noted for vocal variation, clearly tip the burden of proof onto those
maintaining a single species. The sympatry of furcata and glaucops on
Hispaniola clearly merits attention as well, although as I understand it, there
are no genetic data for it. However, the documentation of two or more recently
extinct species of Tyto in the Antilles indicates an appreciable
radiation of barn owls in the Caribbean, of which glaucops and the subspecies
of it, if not themselves genetically distinct, represent the last survivors of
this radiation.”
Comments
from Claramunt:
“YES. As pointed out by others, the situation is complicated, and
there is some critical information missing, in particular, information from
nuclear markers is muted, and it is not clear how the phenotypes sort with
respect to the genetic data. However, accepting that the major mtDNA clades
represent lineages, the study shows that furcata is sister of
Hispaniolan T. glaucops (including punctatissima), not of T.
alba, and knowing that, we should not retain furcata and alba
together to the exclusion of glaucops. Otherwise, relationships would be
misrepresented.
“Furthermore, If T. glaucops and T. furcata, being
so similar in plumage and vocalizations and with 2 million years of divergence,
are already reproductively isolated, that suggests that T. furcata and T.
alba, also similar in plumage and vocalization but with 4 million years of
divergence, may also be reproductively isolated. What do we know about how
these owls maintain reproductive isolation (or not)? Similarities in vocalizations
doesn’t seem to matter much. So, I think that the genetic evidence on actual
relationships trumps speculations about reproductive compatibility in this
case.”
Comments
from Schulenberg:
“As it happens, I visited both Europe and Africa before I ever set foot in
South America. So within just a few years, my birding horizons expanded from
one continent to two continents, three continents, four. Right away I began
keeping track of the species that I had seen on multiple continents. But over
time, those lists began to shrink, not expand, due to taxonomic reassessments.
In a few cases this happened even for species that breed only at high latitudes
(e.g., Velvet Scoter Melanitta fusca and White-winged Scoter Melanitta
deglandi). But the more general trend was of splits that affected species
that were widespread at mid-latitudes, such as Little Tern Sternula
albifrons and Least Tern Sternula antillarum; Kentish Plover Charadrius
alexandrinus and Snowy Plover Charadrius nivosus; Eurasian Magpie Pica
pica and Black-billed Magpie Pica hudsonia;
Eurasian Wren Troglodytes troglodytes, Pacific
Wren Troglodytes pacificus, and Winter Wren Troglodytes hiemalis;
Eurasian Treecreeper Certhia familiaris and Brown Creeper Certhia
americana; and perhaps others that I've forgotten. Viewed in that light,
the possibility that Old and New World barn owls might be separate species does
not seem at all outlandish to me, indeed it seems more likely than the odds
that they're conspecific. So it's well worth keeping an open mind about this
prospect, of course while also searching for additional evidence that would
bear on the question.
“Van
kicked things off by comparing audio recordings of barn owls from various parts
of the Old and New worlds, and came away unimpressed. Dan has a good point,
however, that comparing screeches within the Tyto alba complex only gets
one so far: perhaps one needs a yardstick from elsewhere in the genus for
calibration. To me a broader question here is how well do we understand the
functions of barn owl screeches, and how certain we can be that we're comparing
homologous vocalizations?: these may be a lot trickier to evaluate than, say,
pygmy-owl songs.
“So
one needs a truly deep dive into barn owl vocalizations. Therefore I applaud
Nacho for again calling attention to the discussion in Undiscovered Owls. To
amplify on the piece that Nacho quoted, Robb and The Sound Approach (RATSA)
define the 'perennial screech' as, at least in Europe, 'the ... sound we hear
most often throughout the year'. This is contrasted to a 'courtship screech',
which is seasonal ('peaks during the weeks leading up to egg-laying'), usually
is longer than the courtship screech, usually is given when perched (not in
flight), and differs in a few other parameters (pace, smoother rise in pitch
and volume, etc.). RATSA's take is that the courtship screech is lacking in
American populations. Perhaps the clicking call of North American birds –
something not given by European barn owls - is a homologous vocalization,
perhaps not. In any event, RATSA identify what seem to be significant
differences in the vocal repertoire between American and European members of
the barn owl complex. As noted by Nacho, someone brought this up when this
proposal was before NACC. (I don't know who that was, by the way, but perhaps
Jon Dunn?) I also don't know how much discussion this generated within NACC,
but disappointingly little is reflected in the comments posted online. To me, these vocal
differences are pretty compelling evidence for a split. It's fine to disagree,
if anyone can articulate why you don't find this to be convincing; but simply
ignoring RATSA's take on barn owl vocalizations should not be an option.
“As
far as issues of paraphyly are concerned, discussion to date has focused
mostly, and narrowly, on Tyto glaucops. As background, keep in mind that
early North American taxonomists had no trouble in recognizing New World barn
owls as a separate species from Old World Tyto alba, and also recognized
multiple species in the New World, including Tyto glaucops (e.g. Ridgway
1914, Cory 1918). Hartert (1929) and Peters (1940) later consolidated all taxa
into a single species. In the 1970s North American barn owls colonized
Hispaniola but did not interbreed with resident glaucops, leading to
recognition again of glaucops as a species. So we know that Hartert and
Peters underestimated species diversity among barn owls, but we don't know
whether they were off by just a little, or by a lot. Among other contenders for
species rank are:
• insularis (St. Vincent, Bequia,
Union, Carriacou and Grenada) and nigrescens
(Dominica): These taxa differ from mainland North American barn owls in many of
the same ways as does glaucops, but don't happen to be sympatric with
any other member of the genus. del Hoyo and Collar (2104) classify these as
subspecies of glaucops, despite the range disjunction. These were
recognized as a separate, polytypic species (Tyto insularis) by Ridgway
and Cory. Tyto insularis also was recognized by Suárez and Olson (2020),
based in part of differences in the skull. There are no genetic data on insularis
or nigrescens, nor any detailed analysis of their vocalizations.
• furcata (Cuba and Jamaica): This population
also seems to be pretty distinctive, with white secondaries and tail. It was not recognized
as a species by Ridgway or Cory, but is by Suárez and Olson (2020), again based
in part on osteological distinctions. Note that Uva
et al. included a sample identified as furcata that clusters within
North American Tyto; the relevant sample, however, appears to be a
sequence pulled from GenBank, with no reported locality information, so its
identification as furcata may be questioned, and may not be verifiable.
At the very least, the lack of precise locality for this sample is consistent
with their Figure 1, shows no samples from either Cuba or Jamaica.
• bargei (Curacao): Recognized
as a species by Ridgway and Cory; also accepted, without comment that I could
find, by Suárez and Olson (2020). Genetically, however, bargei
appears to be nested well within North American barn owls (see Aliabadian et al. 2016).
• punctatissima (Galápagos):
Recognized as a species by Ridgway and Cory. As noted by Van, the limited
genetic data (Uva et al.) suggests that it is not
conspecific with mainland North American barn owls. Steadman (1986) also
recognized punctatissima as a species, based on plumage differences and
its much smaller size; Suárez and Olson (2020) seem to concur.
Even
that is not all. Potentially relevant are a host of extinct taxa:
Tyto maniola (Cuba; Late
Pleistocene): a small species; see Suárez and Olson (2020).
Tyto ostologa (Hispaniola;
Quaternary): a giant barn owl; see Suárez and Olson (2015).
Tyto pollens (Bahamas, Cuba; Quaternary): a giant
barn owl; see Suárez and Olson (2015).
Tyto noeli (Cuba, Jamaica,
Barbuda; Quaternary): a giant barn owl; see Suárez and Olson (2015).
Tyto cravesae (Cuba; Quaternary): a
giant barn owl; see Suárez and Olson (2015).
To
recap, there are two widely recognized extant species in the New World
(mainland barn owls and Tyto glaucops); there is good reason to
recognize a third (punctatissima); there plausibly could be one or more
additional extant species; and there are at least five extinct species of New
World Tyto. We don't have a phylogenetic framework that encompasses all
of these taxa (especially of course for the extinct species). The odds of many
of these species resulting from independent colonizations of Tyto from
the Old World seem low, however. Whether each species in the Caribbean
radiation (extant and extinct) represents an independent colonization from the
mainland, or whether there was some in situ differentiation within the
West Indies, remains an open question. Regardless, clearly there has been a
significant New World radiation of Tyto; and it seems quite odd to me to
continue to try to shoehorn mainland North American populations into Tyto
alba while acknowledging not one but a host of New World island species.
And that's not to mention the vocal evidence, cited above, that strongly
suggests that New and Old World barn owls are separate species. This is not a
case, in other words, where we have contradictory information; instead,
multiple lines of evidence all point in the same direction, towards splitting
the barn owls.
Cory, C. B. 1918. Catalogue of birds of
the Americas. Part II, number 1. Field Museum of Natural
History Zoological Series volume 13, part 2, number 1.
del Hoyo, J., and N.J.
Collar. 2014. HBW and BirdLife International illustrated checklist of the birds
of the world. Volume 1. Lynx Edicions, Barcelona.
Hartert, E. 1929. On various forms of the genus Tyto. Novitates Zoologicae
35: 93–104.
Peters, J. L. 1940. Check-list of birds of
the world. Volume IV.
Harvard University
Press, Cambridge, Massachusetts.
Ridgway, R. 1914. The birds of North and Middle America.
Part VI.
Bulletin of the United
States National Museum 50, part 6.
Steadman, D. W. 1986. Holocene
vertebrate fossils from Isla Floreana, Galápagos. Smithsonian Contributions to
Zoology 413. https://doi.org/10.5479/si.00810282.413
Suárez, W., and S. L.
Olson. 2015. Systematics and distribution of the giant fossil barn owls of the
West Indies (Aves: Strigiformes: Tytonidae). Zootaxa 4020: 533–553. https://doi.org/10.11646/zootaxa.4020.3.7
Suárez, W., and S. L.
Olson. 2020. Systematics and distribution of the living and fossil small barn
owls of the West Indies (Aves: Strigiformes: Tytonidae). Zootaxa 4830: 544-564.
https://doi.org/10.11646/zootaxa.4830.3.4
Comments
from Jaramillo:
“YES. Thanks
to Tom for his extensive notes, as well as Nacho for the information, including
the valuable discussions in “A Sound Approach – Undiscovered Owls.”
“Look, I think that the way Barn
Owls use voice in courtship and territoriality is entirely different than in
typical owls. The screech vocalizations are difficult to compare, and there
seems to be a good amount of individual variation in screeches. Even so, in the
Americas there are at least two screech categories, a wider band one, and a narrower
band one. As well, one screech type ends in an upward inflection, almost like a
final accent. I don’t know what this means; however, you can listen throughout
Tyto, and find that it is difficult to hear much difference in
screeches of clearly different (often radically different looking populations,
some sympatric). Sometimes island populations seem to be the most distinct,
visually and vocally. In any case, the fact that so many species have similar
screeches, seems to me that similarity in screech is not a negative for
assessing species limits. Radically different vocalizations are much easier to
apply in a species limit context. But in short,
Tyto are
different than typical owls, and similarity in vocalizations is not all that
informative.
“The fact that the 'kleak-kleak’ call is present in New World
Tyto, and not in the European Barn Owl is huge. As Nacho points out,
this call is common in the New World, and I think it may be a courtship call as
I hear this voice most often in the breeding season, not so much in the winter.
We have Barn Owls commonly here where I live, and a neighbor has a nest box
that is sometimes used just three doors down, so I get to hear them frequently.
This call type is present in glaucops, and also
in insularis
(Lesser Antilles).
“In short, we have genetic data,
and vocal data that clarifies that American Barn Owl should be separated from
the European. By extension, I would separate the Asian-Australasian one as well
to be consistent. It is actually pretty crazy to think that this widespread
taxon would all be one single species. The more we look at Cosmopolitan
species, the more we realize that they are not a single species, and I am sure
we will have others in the future to contend with – Osprey, Great Egret etc.
“I would go further and suggest we
append this proposal to split punctatissima,
or perhaps I can write up a quickie proposal using much of the
data here to make that happen. I recall reading in one of the
Tyto papers how
they found it unusual that two island taxa from Hispaniola (glaucops)
and Galapagos (punctatissima) would be sisters, but this makes a lot of
sense. The connections between the Galapagos and the Caribbean are multiple,
and not only in birds but seemingly in invertebrates as well. There has been so
much work on speciation within the Galapagos, and evolution of forms in the
archipelago that this Caribbean connection has been somewhat lost in the
shuffle. To me separating the Galapagos taxon is clear cut, although we have
little information on it other than skins and photos. Let me assure you that it
is a radically different looking creature. There will be little to no new data
on voice for this species unfortunately. I have tried to get some, but have
never heard it. Similarly, the Galapagos Short-eared Owl is another which I
have not yet heard or recorded. The fact that the fossil record shows a varied
fauna of Tyto over the years is important. Many taxa in the Caribbean are
lineages which are old, and were likely widespread at one point, essentially
living fossils. The proposal that there may still be a greater diversity of
Tyto in the Caribbean/Galapagos is not unusual, it may even be
expected. Here is one Galapagos voice, and it is actually quite different.
Galapagos
https://www.xeno-canto.org/557834
“For the North American committee
it is a tad more complex in that insularis
is also radically different in look (rusty face, ferruginous
overall appearance), and is certainly allied to the
glaucops/punctatissima clade. It
gives the 'kleak-kleak’ calls, and the single screech
that I have heard and recorded was very different from any Barn Owl I have
heard. It seems to me that certainly this is another species, unless it is lumped
with glaucops. Tom’s
note today got me to find my recordings and photos and upload them to
eBird/Macaulay. I offer these to listen to, as they have some pertinence to glaucops/punctatissima. In short,
there are certainly multiple species within the New World, so I do not
understand why there is a reticence to separate the overarching three world
level clades within this context.
insularis
insularis
screech given from perch – St. Vincent (with photos)
https://ebird.org/checklist/S83671424
'kleak-kleak’ calls – Grenada, two
different versions.
https://ebird.org/checklist/S83671800
https://ebird.org/checklist/S83671803
'kleak-kleak’ calls from California for
comparison
https://ebird.org/checklist/S71415159
Additional
comments from Remsen: “At this point, I believe strongly that the
current proposal be tabled and that a new one be written. Actually, a better approach would be for all
this information to be compiled, synthesized, and published as a short
paper. Further, Alexandre Roulin, the world’s authority on the species complex, has
just written a book on barn owls. Roulin is an ecologist, not a systematist, but he has
published perhaps 100+ papers on geographic variation and function of color in
barn owls; I predict his new book will have data or information relevant to
species limits.
“At this point, if this proposal
passes, it will be based on our cumulative comments, not the proposal itself,
and I instinctively dislike that as a matter of procedure.
“As for the Barn Owl, here’s an overview
of the situation. The widespread
continental populations are surprisingly similar in plumage and weakly
differentiated. My read of the weak
genetic data is that New and Old World populations are much more similar
genetically, at least at nuclear loci, than are phenotypically similar
populations of many Amazonian and Andean birds across river barriers. Presumably, this similarity despite the
distance is maintained by dispersal.
Indeed, the Barn Owl sensu lato is certainly among the most remarkable dispersers
in the world. Vagrant records are
numerous; one even reached South Georgia! The propensity to colonize islands is
strong, and so there are numerous spin-off populations that differ from
continental populations to various degrees, likely depending on time since
arrival. Some of these have diverged to
the degree that recognition as separate species is obvious, others only weakly,
and many in between. With such a
tendency for peripheral isolates and rapid speciation, application of paraphyly
criteria to assigning species limits is problematic when comparing insular to
continental populations. What we have
are continental populations, closely related in terms of neutral loci, pumping
out peripheral isolates that then diverge to varying degrees while gene flow
still occurs, evidently, among continental populations, i.e. a situation that
doesn’t really fit the classic allopatric speciation model much less the
cladistic view of endless, clean dichotomous branching.
“Given what seems to be a large
repertoire of weird sounds, I think extra caution is needed when comparing
recordings. Maybe some of these calls
are diagnostically different between taxa, maybe not. How do we know? Without a rigorous analysis
of within-population and among-population variation of homologous vocalizations,
elevating any taxon to species rank or not based on a few recordings seems
unusually unwise, and we have used this very criterion to reject other
proposals on species limits.
“The NACC proposal did not cite “Robb
and the Sound Approach” likely because of lack of access. I’m not sure if I’ve ever seen a copy. At nearly $200 (i.e. in same price range as
an 800+ page, lavishly illustrated HBW volume), I wonder how many individuals
can afford it. The LSU Library, which
has one of the largest university bird libraries in the world, doesn’t have it
(or the Petrel book); I may try to get it the library to order it, although
like most public university libraries, we have budget problems. Thus, a new proposal would need to present
copies of what Robb actually says so that we can do a standard evaluation of
the statements therein in terms of what data support them.
“Although the worldwide distribution of
the Barn Owl certainly makes it a tempting target for taxonomic reassessment,
that sort of distribution itself does not necessarily demand a change in
species limits. The track record of
dispersal of the Barn Owl is also comparable to that of Falco peregrinus
and Butorides striatus, currently treated as polytypic species
(in addition to Alvaro’s examples). As
shown for the Furnariidae by Claramunt et al. (2012: https://royalsocietypublishing.org/doi/full/10.1098/rspb.2011.1922), high dispersal
ability actually impedes speciation.
“Personally, I will continue to vote NO
not because I have any pre-conceived notions of how many species are within T.
alba sensu lato – in fact, the tidbits of evidence cited herein by Nacho,
Tom, and Alvaro suggest current species-level taxonomy is incorrect -- but
because the evidence presented in the proposal is insufficient and because the critical
new evidence presented in the comments, although suggestive, is a compilation
of anecdotes until assembled and synthesized in a coherent way.”
Comments
from Robbins:
“”As Van suggested, will hold off for now on voting on this proposal until a
new proposal is presented.”
Comments
from Pacheco:
“I am convinced from the extensive approach that the
treatment of Tyto alba into three species is the best way to reflect the
evolutionary history of the complex.”