Proposal (922) to South
American Classification Committee
Treat Turdus daguae as a
separate species from Turdus assimilis
Effect on SACC: This would treat the subspecies daguae, our
sole representative of otherwise Middle American Turdus assimilis as a
separate species. Note: this is a slightly modified version of
a proposal submitted to NACC.
Background: This taxon
is currently treated by NACC and SACC as a subspecies of Turdus assimilis, but as a
result of anecdotal comments, NACC and Dickinson & Christidis (2014)
treated it as a subspecies “group”.
Here’s the text from AOU (1998):
“Groups: T. assimilis
[White-throated Thrush] and T. daguae Berlepsch, 1897 [Dagua Thrush]. Turdus
assimilis and the South American T .albicollis Vieillot, 1818
[White-necked Thrush], constitute a superspecies (Sibley and Monroe 1990). Many authors (e.g., Wetmore 1957, Wetmore et
al. 1984, Ripley in Mayr and Paynter 1964) consider them conspecific but see
Monroe (1968) and Ridgely and Tudor (1989).”
I included the part
about albicollis to explain why some older literature treats daguae
under Middle American Turdus albicollis; also, albicollis becomes
part of the problem, as you’ll see below.
Here is our SACC Note
on this:
12a. The subspecies daguae of the
Chocó region has been treated as a subspecies of T. assimilis, and most
recent authors have treated them as conspecific (e.g., Hellmayr 1934, Ripley
1964, Meyer de Schauensee 1970, Ridgely & Tudor 1989, AOU 1983, 1998, Sibley
& Monroe 1990, Clement 2000, Dickinson & Christidis 2014). Ridgely & Greenfield (2001), however,
considered daguae to be a separate species, in part because its voice
resembles that of T. albicollis more than that of T. assimilis. Nuñez-Zapata et al. (2016) presented evidence
that daguae should be treated as a separate species from Turdus
assimilis. Del Hoyo & Collar
(2016) treated as a subspecies of T. albicollis, not T. assimilis,
based on Boesman’s (2016) assessment of songs. SACC proposal badly
needed.
History of taxonomic treatments: Berlepsch
described daguae as a species in Turdus in 1897 from lowland
western Colombia, with the Río Dagua as part of the type locality, which is
south of Buenaventura and NW of Cali (dpto. Cauca). Ridgway (1907) did not mention the taxon, so
I assume it was not recorded in Panama until sometime after 1907. However, Hellmayr (1911; PZSL), Bangs &
Barbour (1922; Bull. MCZ), and Chapman (1926: Birds of Ecuador) soon treated daguae
as a subspecies of Turdus assimilis (at that time known as Turdus tristis).
Hellmayr
(1934) treated it as a subspecies of T.
assimilis with the following
explicit rationale:
“Turdus assimilis daguae
Berlepsch: Differs from T. a. cnephosa [the adjacent subspecies in
central Panama] in smaller size, shorter bill, much darker (bister brown) upper
parts, and very much darker, nearly sepia brown color of the chest, sides, and
flanks. ….
“In coloration, this race
comes nearest to T. a. rubicundus, but is still much more intensely
colored. Although its much smaller dimensions and its shorter, entirely dusky
bill serve to distinguish it without difficulty, yet the close similarity to
the west Guatemalan form seems to afford sufficient evidence for its
association with the assimilis group, which, as suggested by Miller and
Griscom, may ultimately prove to be conspecific with albicollis.”
This treatment was followed by essentially all subsequent authors,
including Ripley in “Peters” (1964), Meyer de Schauensee (1966), and the AOU (1983),
which stated: “The populations of T. assimilis from eastern
Panama (eastern Darien) south to Ecuador are sometimes considered a distinct
species, T. daguae Berlepsch, 1897 [Dagua Robin].” I’m actually uncertain where this statement
“sometimes” came from because I can’t find a treatment from the 1900s on that
did treat it as a separate species. [Anecdote: the frustration that hundreds
of examples like this in AOU 1983 caused me to start lobbying when I joined the
Committee in 1984 for citations for all such statements in future AOU
Checklists; Burt Monroe, who wrote almost all those Notes in the 1983 Checklist
often could not remember the source of many of the statements].
Subsequently, Ridgely & Tudor (1989) treated daguae as a subspecies of T.
assimilis and mentioned only that it was not well known. Clement (2000; Thrushes; Princeton U. Press)
listed it as the southern subspecies of the 10 that he recognized for T.
assimilis; he described the plumage differences but did not illustrate it
separately.
Then, Ridgely &
Greenfield with the collaboration of Robbins and Coopmans (2001; The Birds of
Ecuador Vol. 1) treated daguae as a separate species (Dagua Thrush) from
T. assimilis, with the following text --- note that no actual data are
presented:
“Daguae has
usually been treated as a subspecies of T. assimilis …. Now that
more information is available regarding the voice of daguae – it is distinctly
different from that of T.
assimilis of Middle America – we consider it more appropriate to treat T. daguae as a separate
monotypic species, differing not only in voice but also in several
morphological features. Daguae’s
voice actually more closely resembles that of cis-Andean T. albicollis,
suggesting that daguae may be more closely related to that species.”
Collar in HBW (2005) not only
treated them as conspecific but also treated all of assimilis as
conspecific with South American T. albicollis. He mentioned that daguae had been
proposed as a separate species. Here is
the relevant section of the plate:

Dickinson & Christidis (2014) treated daguae as
conspecific with T. assimilis but placed daguae in its own
subspecies group, as per AOU (1998), and cited Ridgely & Greenfield for the
possible split. Angehr and Dean (2010;
The Birds of Panama. A Field Guide) treated them as conspecific; their range
maps nicely illustrate the substantial gap in their distributions in the
lowlands of central Panama
Del Hoyo & Collar (2016) treated daguae as conspecific
with …. drumroll … not T. assimilis but with cis-Andean T. albicollis. This was based on Boesman (2016), who
actually used the Collar (2005; HBW) species limits, i.e. broadly defined T.
albicollis, not the two species treatment in Del Hoyo & Collar
(2016). His analysis, however, shows
that T. assimilis is clearly different from T. albicollis in song
features, and that the song of daguae is actually difficult to
distinguish from the eastern group of T. albicollis subspecies. Although Boesman would be the first to tell
you that more in-depth analysis is needed, he does establish that if voice is a
reliable indicator, then daguae belongs with T. albicollis. Recall also that Ridgely & Greenfield
(2001) noted the similarity of daguae song to that of albicollis. That trans-Andean Chocó and cis-Andean
populations are sisters is a common biogeographic pattern in Neotropical birds.
Genetic data
As for genetic data, as argued in previous proposals,
I think they are of dubious value for determining taxon rank of allopatric
populations, although perhaps our best estimates of divergence times.
The only study that specifically addressed the daguae
issue was that of Núñez-Zapata et al. (2016), which focused on the T.
assimilis group. Using 2
mitochondrial genes (cyt-b, ND2) and 25 individuals, their tree shows that daguae
is strongly separated, with strong support, from the other subspecies and
populations of assimilis:
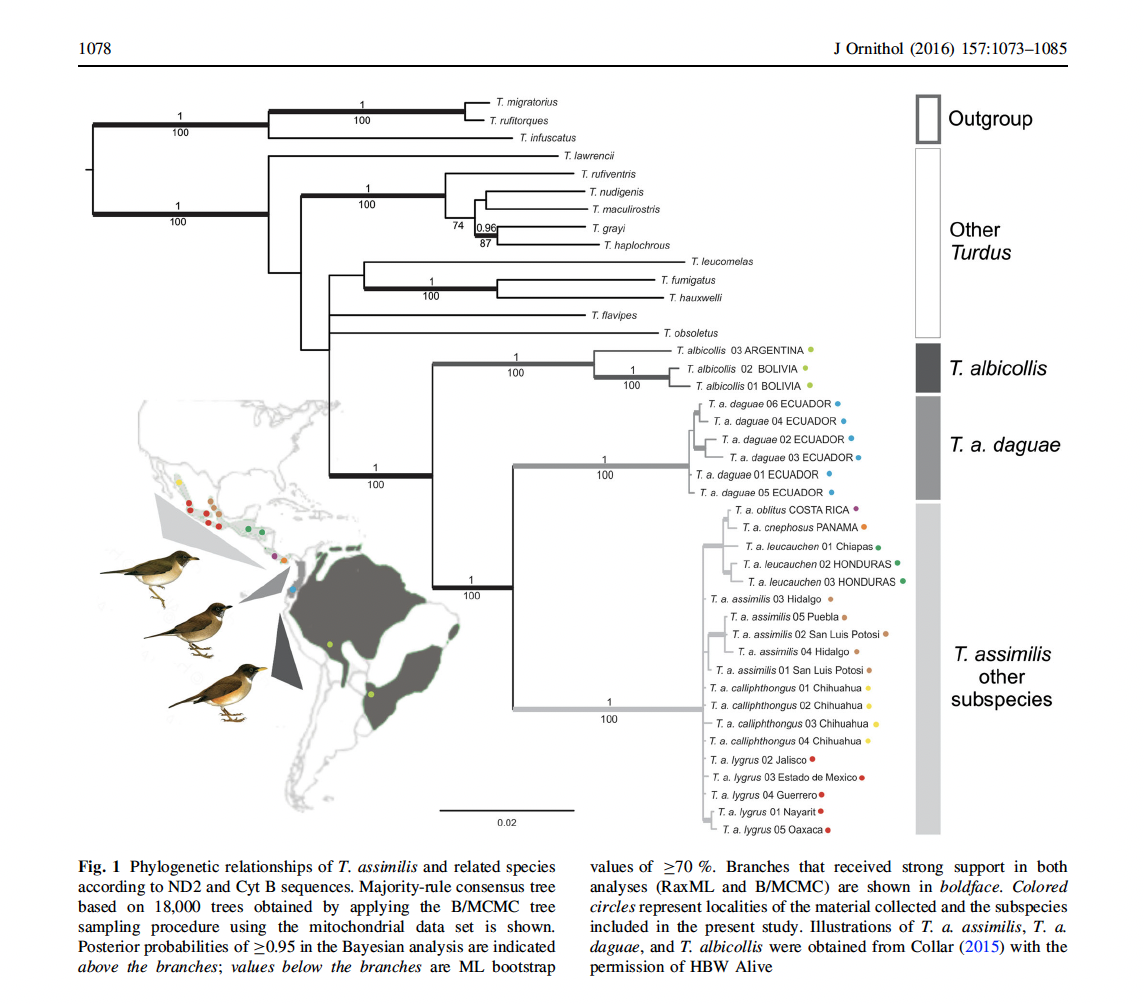
Of interest is that daguae is sister to assimilis,
not the albicollis group, which provides evidence against inclusion of daguae
in T. albicollis as in Del Hoyo and Collar (2016). However, this result could be a “gene
tree/species tree” or ILS problem given the limited gene sampling, and so
hopefully subsequent studies will include nuclear or genomic data. Meanwhile, this places burden-of-proof, in my
opinion on moving daguae to albicollis.
Núñez-Zapata et al. (2016) considered the genetic
data as indicating species rank for daguae based on genetic distance and
reciprocal monophyly. However, they
themselves noted the problems with using comparative genetic distances. Also, with N=6 daguae samples, all
from three localities in NW Ecuador near the southern extreme of its range, a
claim of reciprocal monophyly seems premature.
All such claims are one additional sample away from being refuted. (Would someone please write a paper on the
problems with “reciprocal monophyly” with respect to N and geographic sampling;
certainly, some minimum number of specimens would seem required to make such a
claim, depending on genes sampled, as well as some consideration of the
geography of sampling given that the probability of detecting shared alleles should
decrease to some degree with distance from the former contact zone.)
Batista et al. (2020) used UCEs in their
broad study of Turdus but unfortunately did not include daguae.
Discussion: Voice is the key
indicator of species limits in Turdus and relatives, as outlined in my
proposal on Turdus [m.] confinis. Between xeno-canto and Macaulay there are a
sufficiently large number of recordings from throughout the range of the assimilis-albicollis
group that someone could pick up where Boesman left off and do a formal
analysis of songs and calls to analyze species limits. Until that is done, I do not see how we can
change the status quo. On the one hand,
a preliminary inspection of songs suggests daguae is closer to albicollis
than to where we have it at present, in contrast to a genetic data set that
suggests that it is close to assimilis.
The weaknesses in both analyses make it unwise, in my opinion, to change
current classification for now, either in terms of taxon rank or relationships.
Recommendation: Too much uncertainty
remains, in my view, to make any changes, and so I recommend a NO on this one.
Note
on English names: Dagua Thrush has a track record and is the
only English name associated with the taxon.
Although Río Dagua was an important early collecting locality, it is
nonetheless a pretty obscure river in the greater scheme of things. Whether it’s worth changing to something like
Choco Thrush is at least worth considering.
This taxon’s range corresponds almost perfectly to the Chocó
biogeographic region and its plumage, darkest brown of any on the assimilis-albicollis
group, also reflects a prevailing Gloger’s Rule color trend shown by taxa
endemic to the region. On the other
hand, there are already 9 “Choco Somethings”, so at least Dagua Thrush is
novel.
Van Remsen,
October 2021
Comments
from Robbins: “NO. Without
further analyses of both genetic and vocal characters it would be premature to
treat daguae as a separate species.”
Comments from Lane: “NO. It is frustrating when an
otherwise good study makes one big blunder that severely weakens its punchline.
In Nuñez-Zapata et al (2916), it was the choice of taxa from Turdus albicollis
to include in the study. The samples of albicollis they used were one of
nominate from the Atlantic Forest and two of contemptus from La Paz,
Bolivia. Lowland Amazonian phaeopygius was not included, and this is the
taxon that sounds most like daguae from what I can tell. Furthermore,
whereas albicollis and contemptus are very similar in voice and plumage,
they differ strongly from phaeopygius (to the point that I'd say it's a
no-brainer that the former two should be separated from the latter at the
species level... the question is: where to fit the remaining northern South
American taxa?) the latter is the population that could well be sister to daguae.
The oversight of not including phaeopygius in the study badly damages
the conclusion that daguae should be separated as its own species.
Comments from Pacheco: “NO. Until a comparative vocal
analysis of assimilis/daguae/albicollis with appropriate
geographic coverage is done, I consider it equally premature to alter the
status quo.”
Comments from Claramunt: “YES. Evidence of
reciprocal monophyly is not strong but combined with clear phenotypic
differences in plumage and song, I see a strong case for separating daguae
from assimilis. I totally agree with Dan that the big picture may be
more complex and Amazonian albicollis may be closer to daguae
than to the true albicollis from the south. But I think separating daguae
is a step forward. Once we figure out the affinities of Amazonian albicollis
we can revisit this problem, but I think we don’t need to wait until we know it
all to improve our taxonomy of this complex.”
Comments from Areta: “YES. Nuñez-Zapata et al. (2016) have
provided convincing evidence that daguae is not part of albicollis
(I applaud that they have sampled nominate albicollis), and even if
their dataset is only of mitochondrial DNA it seems highly unlikely that this
pattern will be caused through introgression. Certainly more sampling of ssp.
of albicollis and a proper analysis of vocalizations can shed more
light. But the burden of proof is now on those wanting to treat daguae
as a ssp. of albicollis or of assimilis. Vocally, daguae
sounds different to any ssp. in the albicollis group (at least to my
ear), and the genetic divergence from assimilis (0.051; 1.35 Ma ago
[0.95 to 2.08]) seems to place it within expected distances between good Turdus
species. Biogeographically, it also makes sense to have this Chocó species
being more related to a Central American counterpart. So, although evidence is
not perfect, I prefer to treat daguae as a species on its own.
“Some
bits of information are still missing, but I think a fairly strong case can be
made to recognize Turdus
daguae for the following reasons: 1) Reciprocal monophyly despite
extensive sampling of assimilis,
including geographically distant samples [unfortunately, there are no samples
of daguae closer to the range of the southern subspecies of assimilis, but I don´t think
this will change the picture given the degree of divergence] and 2) distinctive
vocalizations: the song of albicollis
is remarkably constant across its geography (at least to my ears) and I have
never had trouble in identifying this species from Venezuela through NW and NE
Argentina and Brazil, this holds true to the east of the Andes in Ecuador (T. albicollis spodiolaemus),
differing remarkably from the higher-pitched, slower and differently patterned
song of daguae to
the west of the Andes in Ecuador. In this subjective and express analysis, I
agree with Boesman in that songs of assimilis
are the most distinctive, but daguae
seems to me consistently different from albicollis
as well; a thing that always bothers me on these HBW vocal assessments is their
lack of attention to detail and the impossibility of tracking down the
recordings used, which seriously undermines their use as a source of arguments.
I wonder why the apparently distinctive ssp. atrotinctus
was not sampled in the genetic analyses, it does look remarkably different in
plumage! Finally, it is clear that more thorough sampling of albicollis is needed
(sampling in Nuñez-Zapata et al. 2016 is very poor, and samples come only from
the southern range of albicollis),
but I don´t think that this will change the placement of daguae or the degree of
differentiation with respect to assimilis
(note that ssp. phaeopygus
and spodiolaemus
of albicollis in
Batista et al. (2020) are found as sister to assimilis,
which is relevant to the status of daguae).
Comments from Bonaccorso: “YES. Although the genetic
evidence is only mitochondrial, the differentiation is tremendous compared to
all other forms within T. assimilis, and I
agree with Nacho that it is very unlikely that the pattern is contradicted by
additional genetic evidence. Even though the T. a. daguae samples
are from Ecuador, I would not think sampling specimens from northern Colombia
would make a huge difference. I am no expert on vocalizations, but the songs of
T. a. daguae from Xenocanto sound
very different to the forms from Panama and Costa Rica. It is also interesting
that Xenocanto is treating T. daguae separately
from T. assimilis (I guess that is just IOC
taxonomy, but it makes sense to me). It seems that plumage color is not a very
good indicator of species status in this group of very similar taxa. Still,
that is what one would expect from a process of “cryptic” speciation, right?
Finally, this split makes perfect biogeographic sense.”
Comments from Stiles: “NO. Although I tend to agree
with Nacho and Santiago, I agree that there is still sufficient uncertainty
regarding both vocal and genetic data to reach a definitive conclusion. No
genetic data from the albicollis populations closest to daguae,
and while I suspect that tissue samples from Colombia might exist (Check
with Daniel Cadena and Andrés Cuervo). If so, these should be sequenced.
Likewise, need recordings of T. assimilis phaeopygus … so, NO pending
additional data.”
Comments
from Jaramillo:
“YES – Sure, more data are necessary, and some things are unclear. But it seems
to me that as Elisa, Nacho, and Santiago comment, there is enough here to
change the status quo. The status quo appears to be off; we may not quite know
how it is off, but I would rather make this change and adjust at a later date
than not do anything given the data we have.”