Proposal (931) to South
American Classification Committee
Recognize Haematopus bachmani (Black
Oystercatcher) as a subspecies of H. ater
(Blackish Oystercatcher)
Note from Remsen: This is a proposal
submitted to NACC that Terry Chesser asked me to submit here also to SACC to
coordinate our decisions.
Effect on
SACC:
This would lump the Black Oystercatcher (Haematopus bachmani) of Pacific-coast
North America with the Blackish Oystercatcher (H. ater) of South America, considering the former as a subspecies
of the latter, and thus would expand dramatically the range of the SACC
species.
Background:
We are revisiting this species limits issue in
association with the effort to harmonize world lists, and treating H. bachmani as a separate species is a
point of disagreement.
Oystercatcher taxonomy has been an ongoing
challenge because members of the family are morphologically quite conservative.
The two general plumage types, black and pied, tend to correspond to rocky
versus soft shoreline specialization, respectively (Jehl 1985, Hockey 1996).
H.
bachmani
has been recognized as a species by the Check-list
in every edition since the first. The ranges of H. bachmani and H. ater
are entirely allopatric, so the most appropriate way to determine species
limits in this case is to infer them using the classic yardstick comparative
method (although it has been applied infrequently and not with the depth one
might hope for today; Murphy 1925, del Hoyo and Collar 2014). Both taxa occur
in sympatry with the more widespread H.
palliatus (American Oystercatcher), and both hybridize with this pied form
(ater and bachmani are of course of the black plumage type, as their English
names indicate; Jehl 1985, Hockey 1996).
The taxonomic notes provided in the 6th
and 7th editions (AOU 1983, 1998) mentioned that some authors had
considered H. bachmani and H. palliatus (American Oystercatcher) to
be conspecific; they have a hybrid zone of ~480 km in width in Baja California
(Jehl 1985). It is not apparent whether the relationship between bachmani and ater has been evaluated by NACC before.
Murphy (1925:13-15) elaborated on the
differences and similarities between ater
and bachmani thus: For bachmani: “Juvenal birds closely
resemble the young of H. ater, the
feathers of the upper surface, breast, and flanks being edged with pale tawny
brown. It is interesting that the down of chicks of this species is much darker
than that of H. ater or of any other
American form.” And for ater:
“Superficially resembling H. bachmani,
H. ater is widely separated from all
other oyster-catchers in the form of the bill, the excessive compression of
which approaches that of Rynchops.
The distinctive character of the bill is apparent even in chicks taken from the
egg. Color differences between H. ater
and H. bachmani are much greater
among downy young than among adults. The young of ater are relatively pale, only slightly darker, indeed, than those
of H. palliatus, which they much
resemble. The white area is confined to the breast, instead of covering the
belly and flanks as in palliatus, but
it is far more extensive than in bachmani.”
As Jehl (1985) remarked, although AOU (1983)
and Murphy (1925) recognized H. bachmani
and palliatus as separate species,
most other authors did not at that time. For example, H. bachmani and H. palliatus
have been considered conspecific by Peters (1934), Friedmann et al. (1950), and
Mayr and Short (1970)—all of these considered bachmani and palliatus as
subspecies of H. ostralegus, the
Eurasian Oystercatcher. This situation has changed, however, and most
authorities now recognize palliatus
and bachmani as separate species
(checking via Avibase; https://avibase.bsc-eoc.org). Jehl’s (1985) work
appears to have been effective on this issue.
Peters (1934), while lumping both palliatus and bachmani in ostralegus,
considered H. ater to be a separate
species. Although this work does not provide any reasoning behind its taxonomic
decisions (long a sore point), this could have been based on Murphy (1925) or
been simply inertia; as Hockey (1996) related, in the genus in general allopatric
black forms have been considered species, whereas pied forms are often
considered subspecies.
In his study of the hybrid zone between bachmani and palliatus, Jehl (1985) found assortative mating, a stable (though
480-km wide) hybrid zone (after late-19th and early-20th c.
disruption), and inferred selection against hybrids, leading him to conclude
that the two are valid species. He surmised that the primary mechanism of
reproductive isolation was likely postzygotic, focusing especially on predation
on chicks of mixed pairs in which some would have plumage coloration
inappropriate for their beach color, i.e., through loss of crypsis. Given the
high rates of chick predation in some species (60-85%; Hockey 1996), this seems
plausible.
H. ater also hybridizes with H. palliatus, in Argentina (Jehl 1978).
Jehl (1978) described a single hybrid specimen between ater and leucopodus (the
Magellanic Oystercatcher, also a pied form) that he took in Santa Cruz
Province, Argentina. This latter hybridization event (ater-leucopodus) is uncommon compared to ater-palliatus crossings, which he noted occur in this area of
overlap “with appreciable frequency” (Jehl 1978:346). Both the bachmani-palliatus and ater-palliatus hybrid zones should be
revisited with population genetics studies to determine the degrees of
introgression (given clearly incomplete isolating mechanisms), but I do not
know whether such work is occurring.
New
Information
There is remarkably little modern work
available on Haematopus systematics
or species limits. This is an area ripe for study.
Using mtDNA (COI) barcoding, Hebert et al.
(2004) found that the difference between H.
palliatus and H. bachmani was
remarkably low compared with other North American bird species-level
differences, and they considered that this was consistent with treating them as
a single species.
Senfeld et al. (2020) also examined mtDNA (2835
bp) and found palliatus, ater, and bachmani to be very closely related, with bachmani perhaps being sister to the other two. This clade is quite
distinct from H. leucopodus, the
Magellanic Oystercatcher, another pied form as noted above.
del Hoyo and Collar (2014) lumped bachmani and ater, stating that “Race [sic] bachmani
has normally been considered a separate species, but the two are almost
identical in plumage and voice, apparently differing only in greater depth of
bill of nominate ater. Two subspecies
recognized.” (p. 420). (It is worth contrasting this brief emphasis of
similarities with Murphy’s [1925] emphases on differences quoted above.)
Careful analysis of vocalizations is needed.
Subjectively, listening to some of the recordings on xeno-canto (https://xeno-canto.org) reminds me of the
mtDNA relationships: bachmani, ater, and palliatus are similar; leucopodus
is different. It is perhaps no accident that these similarities and differences
are reflected in the rates of hybridization where the taxa overlap. Future work
is also needed to rigorously quantify morphological similarities and
differences. Murphy’s (1925) evaluations show some disagreement with del Hoyo
and Collar’s (2014) conclusions.
With neither appreciable song nor plumage
differences between ater and bachmani, neither assortative mating nor
the putative postzygotic isolating mechanism of strong plumage color selection
favored by Jehl (1985) would likely be very effective in preventing substantial
hybridization (especially given considerable levels of crossing of both with palliatus). I realize that such
conjectures are rather unsatisfactory, but that is one of the acknowledged
weaknesses of the biological species concept when asking whether allopatric
forms are “different enough” to warrant recognition as full species.
Two broader issues have some relevance when
evaluating this and other cases in which allopatry and the Tobias et al. (2010)
criteria are in play. Although many do not like the Tobias et al. (2010)
criteria, the accumulation of subsequent, independent case studies indicate
that the initial use of these criteria has proven much more often right than
wrong in determining species limits (Tobias et al. 2021; although see Rheindt
and Ng 2021 who came to a different conclusion using a different approach). In
addition, across Aves we are likely over-splitting allopatric taxa at the
species level (see Hudson and Price 2014). These are generalities, probably
both true. That said, each case should be rigorously examined.
Taxonomy
and nomenclature:
H. ater (Vieillot and Oudart
1825; Galerie Oiseaux, II, p. 88, I,
P1. ccxxx) has priority over H. bachmani
(Audubon 1838; Birds of America,
folio edit., IV, P1. ccccxxvII, fig. 1). See Murphy (1925) for discussion of
the history of ater and its priority
for that taxon. Thus, if this proposal is approved, H. bachmani would become H.
ater bachmani. Murphy’s (1925) study of the two supports considering bachmani a valid subspecies if lumped
with ater.
Recommendation:
Based on current evidence, particularly the
strikingly different phenotypes of both bachmani
and ater from palliatus and the noteworthy levels of hybridization with that pied
form, these taxa should be considered a single biological species with two
allopatric subspecies-level populations. I find especially compelling that the
strikingly different phenotypes of bachmani
and palliatus appear to be barely
limiting hybridization in a region of overlap to a level that only some
authorities (us included) consider to be low enough to be full biological
species. Given the remarkably close mtDNA relationships among palliatus, bachmani, and ater, it
seems likely that the phenotypic similarities between the latter allopatric
pair (including vocalizations) would be insufficient to preclude more extensive
hybridization if the two were to come into contact.
The vote is in two parts.
A. Recognize H. bachmani as a subspecies of H.
ater.
If A is approved, then B: Apply
the English name Blackish Oystercatcher to both taxa (as HBW-BirdLife already
does).
Literature
Cited
American
Ornithologists’ Union (AOU) (1983). Check-list
of North American Birds, Sixth edition. American Ornithologists’ Union,
Lawrence, Kansas.
American
Ornithologists’ Union (AOU) (1998). Check-list
of North American Birds, Seventh edition. American Ornithologists’ Union,
Washington, D. C.
del Hoyo,
J., and N. J. Collar. 2014. HBW and BirdLife
International Illustrated Checklist of the Birds of the World. Volume 1:
Non-passerines. Lynx Edicions, Barcelona.
Friedmann,
H., L. Griscom, and R. T. Moore. 1950. Distributional
Check-list of the Birds of Mexico, Part 1. Pacific Coast Avifauna 29.
Hebert,
P.D.N., M. Y. Stoeckle, T. S. Zemlak, and C. M. Francis. 2004. Identification
of birds through DNA barcodes. PLoS Biology. 2:e312.
Hockey, P.
A. R. 1996. Family Haematopodidae (oystercatchers). Pp. 308-325 in Handbook
of the Birds of the World, Vol. 3 (del Hoyo, J., t al., eds.). Lynx
Edicions, Barcelona.
Hudson, E.
J., and T. D. Price. 2014. Pervasive reinforcement and the role of sexual
selection in biological speciation. Journal of Heredity 105:821-833.
Jehl, J.
R., Jr. 1978. A new hybrid oystercatcher from South America, Haematopus leucopodus × H. ater. Condor 80:344-346.
Jehl, J.
R., Jr. 1985. Hybridization and evolution of oystercatchers on the Pacific
coast of Baja California. Ornithological Monographs 36:484-504.
Mayr, E.,
and L. L. Short. 1970. Species taxa of North American birds. Publications of
the Nuttall Ornithological Club 9.
Murphy, R.
C. 1925. Notes on certain species and races of oyster-catchers. American Museum
Novitates 194:1-15.
Peters. J.
L. 1934. Check-list of Birds of the World,
Vol. II. Cambridge, Harvard
University Press.
Rheindt, F.
E., and E. Y. X. Ng. 2021. Avian taxonomy in turmoil: The 7-point rule is
poorly reproducible and may overlook substantial cryptic diversity. Ornithology
138: 1–11.
Senfeld,
T., T. J. Shannon, H. van Grouw, D. M. Paijmans, E. S. Tavares, A. J. Baker, A.
C. Lees, and J. M. Collinson. 2020. Taxonomic status of the extinct Canary
Islands Oystercatcher Haematopus
meadewaldoi. Ibis 162:1068-1074. https://doi.org/10.1111/ibi.12778
Tobias,
J.A., N. Seddon, C. N. Spottiswoode, J. D. Pilgrim, L. D. C. Fishpool, and N.
J. Collar. 2010. Quantitative criteria for species delimitation. Ibis
152:724–746.
Tobias, J.
A., P. F. Donald, R. W. Martin, S. H. M. Butchart, and N. J. Collar. 2021.
Performance of a points-based scoring system for assessing species limits in
birds. Ornithology 138: ukab016
Kevin Winker, February
2022
Comments
from Lane:
“NO. Clearly
there is little room for wide variation within the plumages of Haematopus,
with blackish and pied varieties scattered across the coastlines of the world.
They also seem to show some taxonomic constraint in voice, particularly to the
untrained human ear, but in the present case, I would argue that there are
indeed some clear character differences between ater and bachmani.
The former's excitement duet series begins with a rising rapid chatter (not
typically initiated with long notes) that culminates with falling
multi-syllabic notes that look like "M"s or triple-peaked notes (https://macaulaylibrary.org/asset/203976141). The
single note call is in the .11 to .25 second range. By comparison, H. bachmani
enters its excitement duet series with long notes descending into a rapid
chatter that then rises back into a series of couplets that are separate notes
(not multisyllabic notes) (https://macaulaylibrary.org/asset/137867). The
latter is far more like the excitement duet chatter of H. palliatus,
both North and South American forms, by the way (https://macaulaylibrary.org/asset/63003121, https://macaulaylibrary.org/asset/310733941). The
single note call of H. bachmani is in the .29 to .35 second
range. For comparison, H. palliatus has single note calls in the
.28 to .39 second range. [I confess these are based on only a couple recordings
for each species, but eyeballing several recordings of each vocalization type I
think the patterns are real]. So, yes, I would say there are distinct vocal
characters separating H. bachmani and H. ater,
contra what Winker suggests in the proposal.
“The broad overlap of H. ater and H. palliatus
in South America (from Ecuador south to Tierra del Fuego and then north to
central Argentina) with minimal hybridization (I am unaware of any in Peru,
where both occur along the length of the coast), quite unlike the broad hybrid
zone between H. bachmani and H. palliatus (in the
relatively limited overlap they share in southern California and Baja),
suggests that these vocalization differences may be instrumental in maintaining
segregation between H. ater and H. palliatus
whereas the more similar vocalizations between H. bachmani and H.
palliatus could be a reason for the more readily interbreeding of the
two in Baja. Thus, it could be inferred that voice differences between H.
bachmani and H. ater could be a deterrent to interbreeding
between them should they meet hypothetically. I think it is clear that the
"lumperama" days of the last century glossed over some rather
significant characters that differentiated the pied Haematopus (e.g.,
putting H. palliatus or H. finschii into H. ostralegus
simply for superficial plumage similarity and allopatry) and that we should be
wary of doing the same in this century simply because H. ater and
H. bachmani look similar! I vote NO.”
Comments
from Jaramillo:
“NO. I am confused by this proposal. There are multiple elements here that are
not being given due weight in my opinion, and the data is summarized in a way
that suggests that to lump the two black oystercatchers in the New World is
somehow logical. Here are my issues with the proposal.
1)
DNA
– Hebert et al. 2004
https://journals.plos.org/plosbiology/article/file?id=10.1371/journal.pbio.0020312&type=printable
What
can we take away from this paper specifically related to Oystercatchers? I
think three individuals were sampled if I understand correctly. Which
populations of American Oystercatcher? One or multiple? Perhaps we have a hint
that the lineages are recent, but which lineages exactly? I don’t know that we
can deduce anything relating to the question of how many species are involved
from this paper.
2)
Hybridization
– Two hybrid areas are noted. The dynamics of Black and American hybrids in
Mexico are well studied, although the work is now old. A mechanism is given as
to potential selection against hybrid young, as stated in the proposal. But
note that the difference in habitat between Black vs American (Rock vs Sand)
alone, assuming habitat choice is not learned, would be a barrier to gene flow.
The Argentine area of hybrids between American and Blackish is noted, although
this is not well known, and may not be a true hybrid zone as opposed to an area
where hybrids are sometimes noted? I am not clear.
3)
The
thought that a) Black and American show low genetic divergence and b) American
hybridizes with Black at some points, and Blackish at other areas is implied to
mean that gene flow is relatively unimpeded. Maybe? Maybe not. But then I don’t
understand the conclusion to lump Black and Blackish. Shouldn’t the conclusion
be to lump American and Black? And perhaps ask for more work on Blackish to
decide what to do in the future with that? But see below.
4)
NOW
– what is missing here? Well Blackish (ater) and American are sympatric
for thousands of km in Chile and Peru. There are no hybrids that I know of
there. They act like perfectly fine biological species, and you can see them
adjacent to each other, but in different habitat, commonly throughout this
range.
5)
The
proposal mentions that vocalizations seem to be similar between all three taxa,
and are perhaps likely not important as a barrier to gene flow (see final
sentence of recommendation in the proposal). BUT oystercatchers all sort of
sound similar, some are much more clearly different. But they have a whistled
flight call, shorter flight calls, and the duet of quickly repeated notes.
These demand much more study. Because “subjectively” they are similar, and if
you listen to American vs Blackish (ater) you would conclude this to be
the case, yet these two species are sympatric and share a range of thousands of
km. So, something is keeping them apart. Perhaps it is as simple as habitat? I
don’t know. But something is keeping them apart, we cannot discount that
because voice is similar subjectively that it is not doing something, or
perhaps we have not gotten at the difference in that voice.
6)
There
is an assumption that American itself is one thing. However, when I visit Chile
and then I go to Uruguay, the voices I hear are different. I looked at the East
Coast US birds on xeno-canto as well and those sound more like the Uruguay
birds to me as well. It is subtle and this requires more work. But perhaps it
is a mistake to treat American Oystercatchers as all the same thing,
particularly given that they have this linear range, around the perimeter of
the continent, and that Pacific birds are likely not sharing genes with
Atlantic birds at all or have done so less than Argentine American (palliatus)
does with Blackish (ater)! This all needs more work.
Have
a listen – Chilean birds are lower pitched, less strident. Their crescendo
calls do not tend to start with longer, louder whistles. How quickly these
vocalizations are given varies depending on the excitement state of the birds,
but Atlantic birds sound higher pitched, more strident, often with longer
whistled notes at the start. Other oystercatcher species vary in the structure
of the preliminary notes before the duet, so this might be significant. Note
that duets sometimes sound syncopated, other times they do not. The syncopated,
“tic-toc” type situation is more common in the Pacific. Chilean birds are
slower, US and Uruguayan birds (Atlantic) appear to deliver the duet more
quickly. Obviously more study is needed.
But
this does call into question how you evaluate hybridization of ater
(Blackish) with American, as it only happens in the Atlantic, and not the
Pacific. Why?
Chile
US
Uruguay
7)
Voice
of Blackish (ater) and Black (bachmani) differ. I live in the
range of bachmani and hear them on a weekly basis. When I visit Chile, ater
does not immediately register to me. It is subtle. Weirdly enough to me
Americans sound more like bachmani. The voice I am particularly thinking
about is the flight call, which is a whistle. Here at home bachmani
gives this melancholy and relatively long whistle with an inverted U shape on a
spectrogram. Meanwhile ater is much quicker, without thorough analysis I
would say that ater has a call that is often half the length of the
similar call of bachmani. There is variation. however! To my ear the
difference is enough that I recognize a Blackish (ater) as an
oystercatcher, but Black (bachmani) is not what come to my head when I
hear them. Again, the duet voices sound similar, but I bet they are not if
someone was to analyze clearly. But given that flight calls differ between
these two species, I think we might find some differences in the duet call as
well. Would this preclude hybridization? I don’t know, but how ater
interacts with Pacific palliatus (hybrid zone where two ranges meet), is
entirely different than how bachmani interacts with Pacific palliatus
(no hybridization). That alone suggests the two black species should not be
lumped. It is also making an assumption that all Pacific palliatus are
the same lineage, I do not know if that is the case.
Flight
calls ater – shorter more abrupt.
Flight
calls bachmani – longer, more melancholy.
8)
Finally,
morphology. We are overlooking this bill of ater, it is a monster of a
bill. The expansion of the mandible is such that it can resemble a skimmer!
This bill is adapted to entirely different creatures than the smaller bill of bachmani.
Sure, a bill shape might not preclude gene flow if the two came together. But,
assuming a largely genetic component to this particular bill shape, hybrids if
they happened, well they would neither be expected to be able to handle the
mollusks of Pacific South America, or Pacific North America. That is to say,
that this is not just a slightly large bill, it is an entirely different bill
shape. BTW, I think the same of Ringed Kingfishers in the Temperate zone and
the Tropical zone, they are so different in bill shape that I bet that hybrids
would not do well. But that is another story.
9)
To
me, much more work is needed. There are definitely some young lineages here, an
effect of habitat and ecological divergence, and perhaps an over emphasis on
lumping due to similar plumage of the two black species. I would not be
surprised if they are both offshoots of the American clade that may look
similar due to their habitat, but they may each be more closely related to one
of the American populations than they are to each other. But the fact that in
the Pacific of South America Blackish (ater) is sympatric with one form
of American, well that is enough for me to say, keep them all separate. And
that more work should be done to understand the dynamics within palliatus,
let alone how it relates to the two black species.”
Comments
from Peter Boesman: “Notes on the vocalizations of Black
Oystercatcher Haematopus bachmani and Blackish Oystercatcher Haematopus
ater:
“I
compared the sonograms of the Haematopus species, and most look very
much the same. Only in some cases of largely sympatric breeding areas of two
species of the genus, there seems to be some clear vocal differentiation, e.g.,”
Haematopus
ater vs. H. leucopodus: the latter has mainly
high-pitched squeaky notes in its vocabulary
Haematopus
finschi vs. H. unicolor: the latter has some more nasal
notes in its vocabulary
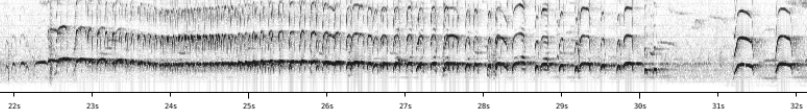
“In the specific case of Haematopus ater and H.
bachmani I think however I have found one difference! In the former
the long piping trill typically uttered during display or interaction
apparently always starts at the lowest frequency after which it rises in pitch
for a while to remain stable afterwards. In the latter it typically starts at a
high pitch after which it drops and rises a bit to remain stable afterwards.
Examples:
H.
ater (XC188260)
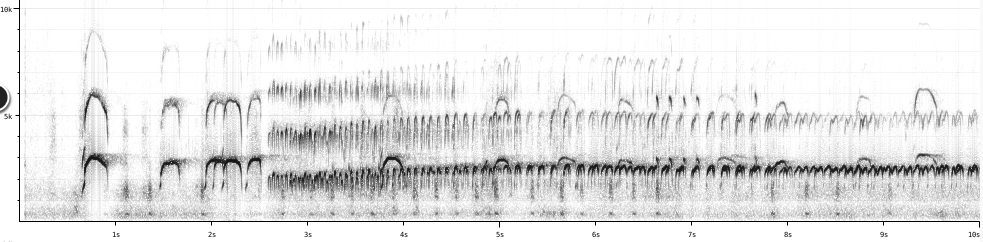
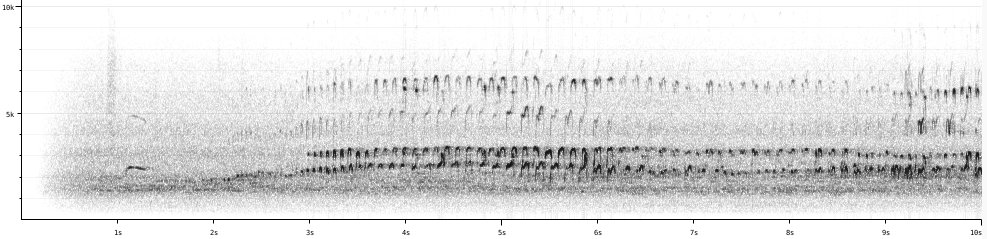
H.
bachmani
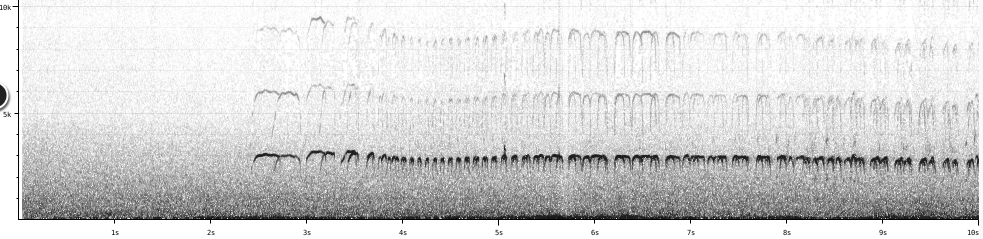
“There
aren’t that many recordings in which the full series is recorded from the very
start, however:
H.
ater
XC:
4 recordings, all have initial rise
ML:
2 recordings, all have initial rise
H.
bachmani
XC:
7 recordings, all showing the initial drop and rise in pitch
ML:
I didn’t listen to all 226 recordings but of the 7 I found having the piping
trill, all showed the initial drop and rise.
“Whether this is an important difference is hard to tell
but given the vocal variation in the genus is very limited, and given it is a
vocalisation used during display and interaction, it may well be considered
important. If it has to be scored for Tobias, one could define the parameter
initial drop or rise in pitch, which would give a score of about 3.
“A
possible second difference is that H. ater starts the
trill with very short notes which gradually increase in duration, whereas bachmani
has initial longer notes which reduce in duration during the drop in
frequency. I said ‘possible’ because one can’t be sure always if the initial
longer notes are from a second bird or not, but it would seem this feature
holds quite well, in which case it could also be given a score of 2-3.”
Comments from Remsen: “NO. The points made
by Dan, Alvaro, and Peter all strongly indicate not only that more work is
needed but also that bachmani and ater are unlikely to be sister
taxa, as suggested even by the weak genetic data in Senfeld
et al. It would seem to me that the
necessary first step in treating bachmani and ater as conspecific
would be to show that they are more closely related than either is to palliatus,
using more than just mtDNA and with broad geographic sampling. Until that necessary condition is met, I don’t
see any point in changing current species limits.”
Comments
from Robbins:
“NO for treating Haematopus bachmani as a
subspecies of H. ater for reasons detailed by all who have commented on
this proposal (as of 2 February 2022). Clearly, more study is needed before
making any taxonomic changes.”
Comments
from Areta:
“NO to H. ater bachmani. Although
there is clearly fertile ground for research, current vocal, plumage, and
morphological differences argue against the lump (thanks Alvaro, Dan, and Peter
for insight into some of these aspects).
“Phylogenetic
information (mostly Senfeld et al. 2019) and data on
hybrids also indicate that considering this case without including H. palliatus in the taxonomic debate is
futile. It is also the case that Murphy (1925:5) reported on the long wings and
large feet of H. ater in comparison to other Haematopus,
especially bachmani: "Of the native American species of Haematopus (H. ostralegus
being excluded) ater has by far the
longest wings and the largest feet, being in these respects at great variance
with the other black oyster-catcher, H.
bachmani."
“Regarding
the Pacific vs. Atlantic forms that Alvaro has mentioned, Murphy (1925: 4)
reads: "From still another geographic placement,
it is noteworthy that the Pacific races of ostralegus
and palliatus (e.g., longirostris, frazari, galapagensis, pitanay) have unmarked primary
quills, while the Atlantic races of both these species (e.g., H. ostralegus ostralegus,
H. palliatus paliatus,
H. p.prati,
H. p. durnfordi)
all share in greater or less degree the peculiar pattern. The rule breaks down
only near Panama, where the West Indian maritime avifauna has so generally
crossed over to the Pacific side". It is clear that
unless H. palliatus is studied at
length and across geography, we won´t have a clear picture of what to do with H. bachmani.
“I
see nothing in the proposal to change the status
quo of recognizing H. bachmani as
separate from H. ater.”
Comments from Pacheco: “NO. Given the informative
considerations made by Dan, Alvaro, and Peter, it is unsuitable to accept the
downgrade of Haematopus bachmani to subspecies.”
Comments
from Claramunt:
“NO. Similarity has never been a good guide for lumping forms into the same
species, and in this case there are clear morphological (bill shape is
diagnostic) and plumage (in juveniles, as pointed out by Murphy 1925)
differences.”
Comments from Bonaccorso: “NO. I think Alvaro´s detailed
reasoning is quite convincing. I vote no until more data and detailed analyses
are presented.”
Comments from Zimmer: “NO, for all of the reasons
nicely summarized by Dan, Alvaro, Peter, and others. I would add that the difference in plumage
ontogeny between ater and bachmani strike me as indicators of
distinctiveness too – juveniles of the two taxa are nearly identical in plumage,
whereas downy chicks differ significantly in color (and the differences in bill
morphology are also already apparent in downy chicks). As others have already elaborated, I think it
is dangerous to include alleged vocal similarity as a rationale for lumping
taxa, in the absence of any kind of quantitative analysis, and based solely
upon a gross qualitative comparison of how these things sound to the human
ear. And, as pointed out by Van and
others, any reevaluation of species limits as regards ater and bachmani, really
needs to include a proper analysis of the relationships of each of those taxa
to palliatus, and with broad
geographic sampling throughout the rather extensive ranges of all three species.”
Comments
from Stiles:
“NO - multiple lines of evidence clearly indicates
that lumping bachmani into ater is not a good option.”