Proposal (956) to South
American Classification Committee
Species
limits and generic placement of Phaeomyias
murina
Our current SACC note reads:
“20. Ridgely
& Tudor (1994) noted that vocal differences suggest that Phaeomyias
murina might consist of more than one species. Ridgely & Greenfield
(2001) considered the subspecies tumbezana (with inflava and maranonica)
of southwestern Ecuador and northwestern Peru to represent a separate species
based on differences in vocalizations.
Rheindt et al. (2008c) found genetic evidence consistent with two
species, and Zucker et al. (2016) found additional evidence for multiple
species within P. murina. SACC proposal badly
needed."
Zucker et al. (2016) provided key material to discuss
species limits in Phaeomyias murina.
What they found is that the Cocos Flycatcher, historically placed in the genus Nesotriccus, is embedded
within Phaeomyias
and that the latter includes 4-5 distinct clades, which might merit species
status. Harvey et al. (2020) recovered a similar phylogenetic structure. Lanyon
(1984) had already shown that Phaeomyias
and Nesotriccus
were closely related, and Rheindt et al. (2008) also provided phylogenetic
information to begin to unfurl this riddle. This is a good example of why “express”
splits should not be done even if obvious and why deep studies are needed:
there are surprises hidden everywhere.
As I see it, we have the following options:
1)
Split P.
tumbezana (with ssp. inflava): tumbezana/inflava and maranonica
are narrowly parapatric, are deeply diverged and differ in vocalizations. Zucker
et al. (2016: 300) wrote:
"Lowland tumbezana and montane populations
matching maranonica in plumage,
voice, and mitochondrial DNA occur within about 10 km of each other on the
lower slopes of the western Andes, where they appear to segregate by habitat
and elevation (Angulo et al., 2012; Schmitt et al., 2013; F. Angulo P., D.
Lane, pers. comm.). Vocal, morphological, and genetic data divergence between tumbezana/inflava and maranonica (including montane Tumbes
populations), combined with their nearly sympatric distributions, suggest the
two merit recognition as separate species. Further work is needed to ascertain
if interbreeding or introgression occurs in this region."
2) Split P.
maranonica: keeping the splits 1 and 2 under a single species P. tumbezana would be
another option, but this is not recommended given the vocal differences (see
Schulenberg et al. 2007), the relatively deep genetic break and the near
parapatry of these taxa.
3) Split P.
incomta (with ssp. eremonoma):
note that the paper by Kroodsma et al. (1987) on the
vocalizations of ridgwayi,
coupled to its divergent morphology (see Sherry 1985, cited in Sherry 1986),
and level of genetic divergence argue against the lump of ridgwayi with incomta/eremonoma.
I am not aware of any formal analysis comparing incomta/eremonoma vs. murina/wagae/ignobilis, but
the differences are striking based on my recordings and field experience with
both in Argentina/Bolivia and Venezuela, backed up also by recordings by others
(notably by P. Schwartz in Venezuela). The calls, diurnal song and dawn songs
are clearly different, even when they share an overall "Phaeomyias"
feel/structure to them.
4) Further splits in P.
murina/wagae/ignobilis: This clade is possibly the most problematic
because there is a relatively deep split that may indicate the existence of two
species. The type of P. murina
is lost (type locality Brazil), and wagae
is from E Peru (type locality Chirimoto), and ssp. ignobilis (type locality Villa Montes, Bolivia)
was lumped with murina
by Fitzpatrick (2004). Therefore, without more work comparing type specimens
and matching those to the clades in Zucker et al. (2016), it is not clear which
name should be applicable to which population, and it looks like a decision should
be made regarding a restricted type locality and possibly a neotype designation
for murina (which
apparently have not been done). I also want to note that at least the southern
populations traditionally ascribed to nominate murina and ignobilis
are highly migratory, and that it is likely that several of the more northern
examples from the Peruvian Amazon (green spots) in the tree of Zucker et al.
(2016) are southern migrants. The problem with “who is who” gets diluted (at
the species level) if one decides not to split these two clades, which is
probably the best course at present given the lack of detailed studies
analyzing their vocalizations, the borderline genetic differences and the mess
with which names to apply.
5)
Phaeomyias vs. Nesotriccus: A final note has to do with the
generic names. Nesotriccus Townsend
1895 has priority over Phaeomyias
Berlepsch 1902 (type species Elainea
incomta). Given the position of Nesotriccus
and the levels of divergence, it seems that all the taxa should be placed in Nesotriccus, which is rather
unfortunate from the standpoint of stability. It seems an inescapable change.
Rheindt
et al. (2008):
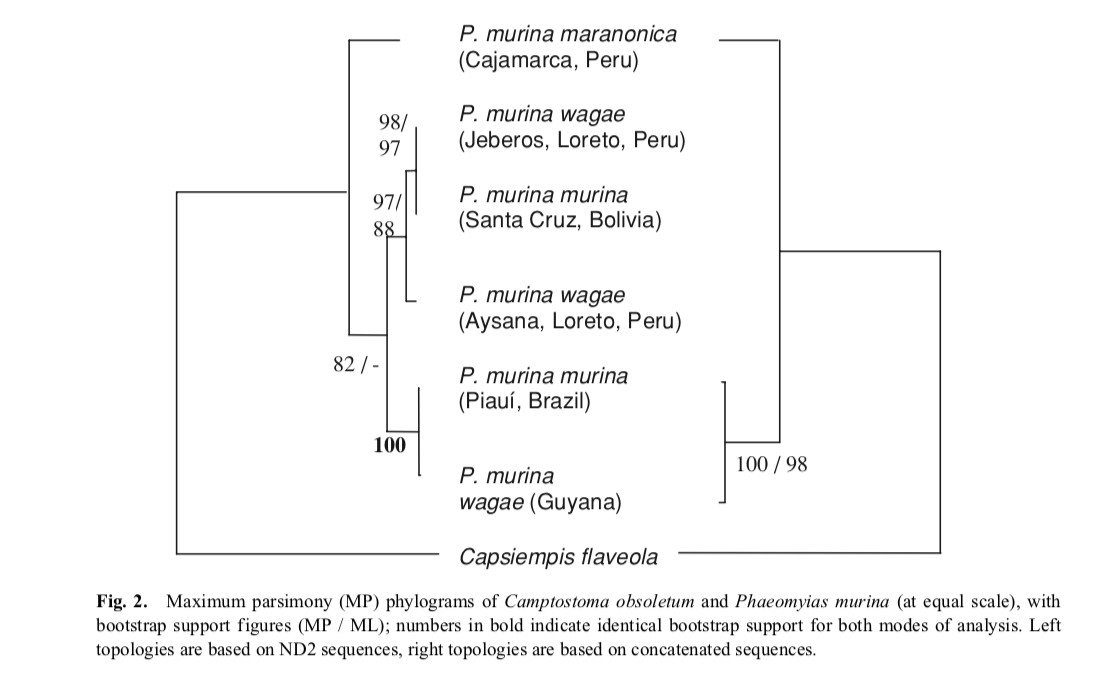
Zucker
et al. (2016):
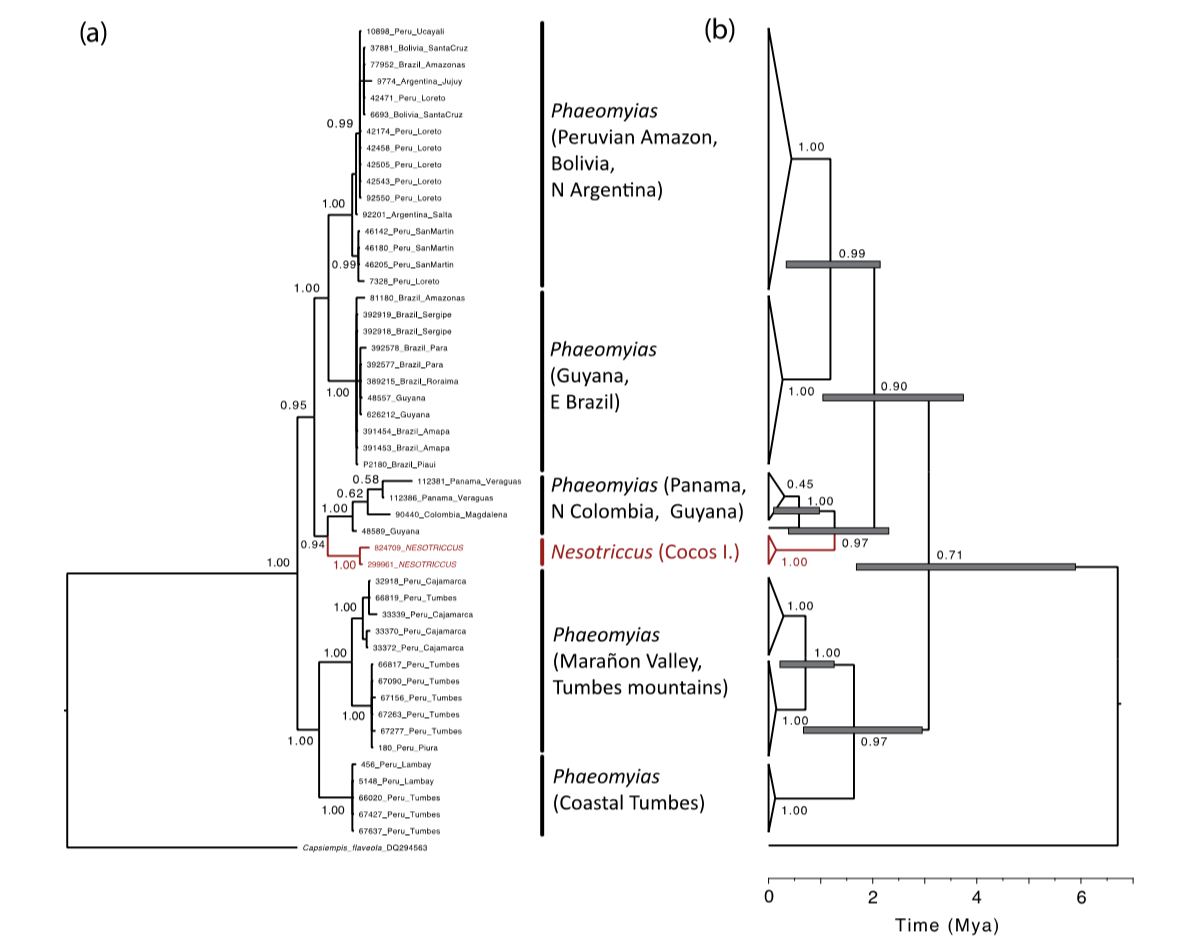
Harvey
et al. (2020):

Note on gender of names: Nesotriccus being masculine (Dickinson & Christidis 2014),
variable epithets need to be corrected, which impacts almost every species and
subspecies names in this group. Only the subspecies wagae is invariable (Dickinson & Christidis 2014). Therefore,
the corrected names would be:
Nesotriccus murinus
Nesotriccus murinus murinus
Nesotriccus murinus wagae
Nesotriccus incomtus
Nesotriccus incomtus incomtus
Nesotriccus incomtus eremonomus
Nesotriccus maranonicus
Nesotriccus tumbezanus
Nesotriccus tumbezanus tumbezanus
Nesotriccus tumbezanus inflavus
Discussion:
I recommend
YES to splits 1, 2 and 3
(i.e., recognize P.
tumbezana, P.
maranonica, and P.
incomta) and YES to keep P.
murina/wagae/ignobilis as a single species until further work
clarifies what is going on there.
The Guyana samples falling in the murina clade or that in the incomta clade might be seen
as problematic by some. However, the ones in the murina clade could pertain to southern migrants
(I am not sure what these specimens look like, and I cannot access the
supplementary material to see whether the collection date makes sense for a
migrant). Regardless of this, Guyana is not the type locality of any of these
birds, and having two nearby samples falling in different clades can also be
interpreted as evidence of two species (much as in tumbezana and maranonica). Also, keeping incomta and murina/wagae/nobilis clades
as one unit creates a somewhat odd paraphyletic species with two distinct vocal
types (i.e., vocalizations of incomta
and the murina/wagae/nobilis
clade differ noticeably, even when no formal analyses exist). I therefore think
that the split 3 is quite straightforward, but what I think is not safe to do
is to perform any more splits in the murina/wagae/nobilis
group; therefore, I recommend a NO on 4.
Also, I recommend a YES on including all species in Nesotriccus.
English names: To be determined in a subsequent proposal
depending on which parts of this one pass.
References:
Angulo, F., Flanagan, J.N.M., Vellinga,
W.-P., Durand, N., 2012. Notes on the birds of Laquipampa Wildlife Refuge,
Lambayeque. Peru. Bull. Br. Ornithol. Club 132, 162–174.
HARVEY, M. G., G. A. BRAVO, S. CLARAMUNT, A. M CUERVO, G. E. DERRYBERRY,
J. BATTILANA, G. F. SEEHOLZER, J. S. MCKAY, B. C. O’MEARA, B. G. FAIRCLOTH, S.
V. EDWARDS, J. PÉREZ-EMÁN, R. G. MOYLE, F. H. SHEDLON, A. ALEIXO, B. T. SMITH,
R. T. CHESSER, L. F. SILVEIRA, J. CRACRAFT, R. T. BRUMFIELD, AND E. P.
DERRYBERRY. The evolution of a tropical
biodiversity hotspot. Science 370:
1343-1348.
Kroodsma, D. E., V. A. Ingallis, T. W.
Sherry, and T. K. Werner. 1987. Songs of
the Cocos Flycatcher: vocal behavior of a suboscine on an isolated oceanic
island.. Condor 89: 75-84
Lanyon, W.E., 1984. The systematic
position of the Cocos Flycatcher. Condor 86, 42–47.
Rheindt, F.E., Norman, J.A., Christidis,
L., 2008. Genetic differentiation across the Andes in two pan-Neotropical
tyrant-flycatcher species. Emu 108, 261–268.
SCHULENBERG, T. S., D. F. STOTZ, D. F. LANE, J. P. O'NEILL, AND T. A.
PARKER III. 2007. Birds of Peru.
Princeton University Press, Princeton, New Jersey, 656 pp.
Schmitt, C.J., Schmitt, D.C., Tiravanti,
J.C., Angulo, F.P., Franke, I., Vallejos, L.M., Pollack, L., Witt, C.C., 2013.
Avifauna of a relict Podocarpus forest in the Cachil Valley, north-west Peru.
Cotinga 35, 17–25.
Sherry, T. W. 1986. Nest, eggs, and reproductive behavior of the
Cocos Flycatcher. Condor 88: 531-532
ZUCKER, M. R., M. G. HARVEY, J. A. OSWALD, A. M. CUERVO,
E. DERRYBERRY, AND R. T. BRUMFIELD.
2016. The Mouse-colored
Tyrannulet (Phaeomyias murina) is a
species complex that includes the Cocos Flycatcher (Nesotriccus ridgwayi), an island form that underwent a population
bottleneck. Molecular Phylogenetics and
Evolution 101: 294–302.
J. I. Areta, February
2023
Comments from Lane: “Well
drat. This proposal basically has rendered a paper I am working on as largely
moot, which is irritating ... not least of which because Tom asked me for
information on this case, and I relayed to him but that I was writing about it
currently. I agree that the species split is clearly necessary, as is the
reason for synonymizing Phaeomyias under Nesotriccus; so, YES to parts 1-3 and 5, and NO to part 4 (recognizing splits within murina/wagae/ignobilis).
Zimmer made a few unusual mistakes when reviewing these taxa, which has
resulted in compounded taxonomic and distributional mistakes through all
subsequent works, and I hope that that topic will remain relevant enough to
warrant me finishing and publishing my work.”
Comments
from Claramunt:
“YES to treat Phaeomyias species under the
genus Nesotriccus, unfortunate but unavoidable. Reluctant YES to 1, 2,
and 3 because the information is not all out and analyzed in a peer reviewed
paper. NO to part 4. Dan: we still want to see your paper.”
Comments
from Stiles:
“A,B-both YES to splits of tumbezana and maranonicus given
genetic differences and probable parapatry, maintaining ridgwayi as
separate; C-YES to separate incomta and eremonoma on genetic
distinctions; D-NO to further splits without further genetic and vocal data; and E-YES to Nesotriccus
for all, its priority is undeniable.”
Comments
from Shaun Peters:
“Normally I
would not get involved in the SACC proposal process, but since I am preparing
some comments to send to both Clements-eBird and IOC regarding their split of Phaeomyias
murina and some other taxonomic decisions (presumably based on a what is
coming from WGAC), I thought I'd share my thoughts with you, which you will
find in the attached pdf* (converted from original word file to reduce file
size). Also attached are the maps from the pdf and Supplementary Table 1 from
Harvey et al listing specimen localities.
“To summarize:
“The proposed
split of
P. murina in to 4 species is essentially based on
genetic data and, in particular, paraphyly. This mainly comes from Zucker et al. (2016), but they only used a single mitochondrial gene (ND2). Harvey et al. (2020), which used UCEs, has limited sampling (6 samples,
no samples from P. m. murina). The trees in these two papers are
broadly similar with the tumbezana and murina groups forming separate clades and Nesotriccus ridgwayi as sister to the incomta group, but not with true paraphyly, but what I call
'pseudo-paraphyly' - this being the case in Harvey et al and also in Zucker et al once the tumbezana clade
is separated off as a separate species. 'Pseudo-paraphyly' does not require a
mandatory split under a BSC.
“Zucker et al. then proposed a split of maranonica (which now includes Andean tumbezana) from lowland tumbezana/inflava, mainly
based on their deep genetic divergence, but also citing differences in
morphology and vocalizations. There do appear to be differences between coastal tumbezana and maranonica, but there are few
available recordings of Andean tumbezana and inflava (the latter being the most distinct taxon in
morphology), and there may well be vocal differences between inflava and lowland tumbezana.
“Harvey et al. then split incomta from murina based on paraphyly but, as previously stated, this is
actually a case of 'pseudo-paraphyly' so is not necessary under BSC. Nacho
mentions differences in vocalizations, but the situation is a little more
complicated than he stated. Listening to vocalizations over the whole range of
the murina group, eastern and southern birds (murina)
generally have rather burry dawn songs and calls with northern birds (incomta)
having clear dawn songs and calls, although some daytime songs from incomta have a burry quality. Interestingly, recordings of wagae resemble incomta in their pure tone, whilst those of ignobilis seem closer to murina than wagae. Thus there may well be three (or four) vocal types within
the murina group. There is also the fact that incomta and murina are
very similar in morphology and it is wagae that
stands apart (see Zimmer, 1941, although based on Dan's comments on the
proposal this may not be the case??).
“Thus, there may
well be more than one species involved within the murina group, but it requires more widespread genetic analysis
(of both nuclear and mitochondrial genes) as well as a detailed comparison of
vocalizations across the whole range of the murina group.”
[* distributed to SACC
members separately]
Comments
from Shaun Peters (now voting for Pacheco): “Since a
split of a broad tumbezana (including maranonica) is not on the table I would have to vote 'No' on Parts 1
and 2. Although lowland tumbezana and maranonica clearly differ vocally, I'm not certain about the vocal
affinities of Andean tumbezana and inflava (more work is needed here). 'No' votes on parts 3 and 4 are
more straightforward for me - more work (how the genetic, morphological and
vocal data marry up) needs doing here, Thus, here are my votes
“Part 1 - NO
Part 2 - NO
Part 3 - NO
Part 4 - NO
Part 5 – YES”
Additional
comments from Areta:
“Shaun, thanks for the diligent analysis shared. I should
have checked for the restricted type locality for murinus; however; even if there might be more splits in the murina-wagae-ignobilis, data are far from satisfactory and nomenclatural problems
should be cleared before proceeding on this front. For example, although murina has been
restricted to a type locality, apparently no neotype has been designated; thus,
flimsy ground upon which to decide.
“Also, bear in mind that the support for the Guyana and Colombia
samples in the Harvey et al tree is so low as to render this a complete
uncertainty, and cannot be interpreted as introgression with any confidence.
So, there is no "pseudo-paraphyly" here.
“After reading Shaun’s comments (and incorporating the caveat that
the Harvey et al. tree cannot be used to discard or confirm paraphyly in the murina
group), the genetic and vocal data are consistent with the treatment that I
advocate: split incomta/eremonoma, split tumbezana, split maranonica,
and leave murinus as one species until the proper studies needed to sort
out their taxonomy are published (it is possible to split the NE and SW clades
as separate species, but I didn´t want to go that far in the proposal, given
the confusion surrounding the distribution of the different taxa, the lack of a
proper vocal analysis, and type-specimen issues). As I mentioned in the
proposal, the likely coexistence of taxa from two different clades in Guyana
provides further support to split incomta/eremonoma from the murinus
group. Now, whether these breed there or not, I don´t know. Maybe Dan has
researched this more in-depth for his paper.
“I think that there are plenty of questions here to be answered,
but I think that the SACC proposal is consistent with the minimum number of
necessary splits that are well-supported by the data."
Comments
from Bonaccorso:
“YES to part 5. The evidence from the broad sampling
of Zucker et al. (2016) on ND2 and the restricted UCEs sample from Harvey et
al. (2020) are consistent with Nesotriccus well nested within
Phaeomyias. If Nesotriccus is an older name, the change of all Phaeomyias of Nesotriccus is adequate.
“NO to the splits (parts 1, 2, 3, 4). The splitting of these taxa
would be based solely on Zucker et al. (2016) ND2 data and their presumable
paraphyly. Such a split would need a more integrative approach with more
genetic data, including potential contact zones and some diagnostic characters,
either morphological or vocal.”
Comments from Robbins: “The combination of genetic and
vocal data do support the minimum splits that Nacho has proposed, thus I vote
for acceptance of 1,2,3, and of course, # 5, using Nesotriccus for all
these taxa.
“With regard to whether birds breed in Guyana, samples that we
collected during March in the Rupununi do breed there (see Appendix 1 in
Robbins et al. (2004. Avifauna of the Guyana Southern Rupununi, with
comparisons to other savannas of northern South America. Ornitologia
Neotropical 15:173-200). For example, I
recorded persistently singing birds (ML 145030, 145010) on the same day/site
that birds were collected that had enlarged testes, e.g., 7 x 4 mm (KU 90771). With regard to the two LSU Guyana samples in
Zucker et al. (2016), thanks to Steve Cardiff, a photo is attached below of
those specimens (collected by Santiago). Note how strikingly different those
two specimens are, even though both are adult females with similar gonadal
data, taken at the same locality within two days of each other (during the
austral winter). So, I suspect one is a
migrant and the other a resident. The
paler bird is more consistent with material that we have collected throughout
the year in Guyana.”
“To add a bit of perspective, three specimens (KU 90771, 90884-5)
taken in the southern Rupununi during late March to mid-April are
indistinguishable from a specimen with enlarged testes (6 x 3 mm) taken on 31
October (KU 96872) from Jujuy, Argentina! Furthermore, in Birds of the World, which has
already split the murina complex into multiple species, states the two
subspecies of the Northern Mouse-colored Tyrannulet (Nesotriccus incomta)
differ in that Central American N. i. eremonoma is distinguished from
nominate by having the wing coverts edged dull buff. The aforementioned March-April birds from
Guyana have the wing coverts edged dull buff.
“So, I would submit that delineation of subspecies using plumage
would seem highly problematic: voice and genetic data should define taxa in
this complex.
“Clearly, more in-depth study
is needed, but I think it is a major step forward in recognizing the proposed
splits in this proposal.”


Comments
from Zimmer:
“This is definitely a messy taxonomic and nomenclatural case, and one for which
we don’t have enough data to completely untangle. However, I think there is enough evidence for
us to move things forward on some fronts, so, here goes: YES to splits 1, 2 & 3, in
spite of some unresolved issues regarding vocal variation within tumbezana
(coastal versus Andean) and how that relates to both inflava and maranonica. I think it is a step forward to make these
splits to at least begin to address some of the vocal and genetic
variation. But, like Nacho, I don’t
think we know enough about what’s going on to justify any further splitting at
this time, within the murina/wagae/nobilis group, so NO on 4. Also, a clear YES on 5, since Nesotriccus
is clearly embedded within Phaeomyias, and the priority of Nesotriccus
is undisputed.
Additional
comments from Lane:
“Thanks to some queries from Niels Krabbe on voices
of some Ecuadorian populations of this complex, I have reviewed recordings in
both Xeno-canto and Macaulay Library and found that it seems that both have
made a real hash of the application of taxonomic names to populations. Macaulay
has already instituted the split of N. incomtus from N. murinus, but many
of the recordings placed in the former sound more to me like wagae, which is placed in the latter, and which is the
cis-Andean form I know best. As Nacho lays out, the type locality of incomtus is Cartagena,
and there are not a lot of recordings from around there for me to feel like I
really have a grasp of what that taxon sounds like (I can't find any real dawn
song from around Cartagena, for example). My gut feeling is that the name incomtus is best
applied to birds from northern Colombian lowlands east along the Venezuelan
coast (possibly in the northernmost edge of the Llanos as well?), and,
presumably, into Guyana (as per the Zucker et al. phylogeny), but I don't hear
recordings from Guyana that sound like those from Colombia. To my ear, many of
the dawn songs available in ML from Venezuela and Guyana sound very much like
the waga
song I know from Peru (see here for a near
topotypical example: https://macaulaylibrary.org/asset/234404). Indeed,
nearly all the highland birds on XC from central Colombia and Ecuador seem to
sound much more like wagae, but with
higher-pitched voices. This suggests that the
incomtus group is
far more range-restricted than it seems to have been thought (at least based on
the maps available on XC and eBird/Birds of the World... which may or may not
be of any value in this discussion anyway), but more importantly, unless each
listener is carefully combing the recordings for topotypical examples and
working outward from there, they will be completely swamped by misinformation
as to the voice characters of each group. I hear much variation within all
these murinus/incomtus
populations, including what appears to be several unrecognized
taxa (the highland birds from Colombia and Ecuador, and then birds from NE
Brazil seem fairly distinctive compared to other cis-Andean populations. That
last tidbit calls into the light the proper application of the name
murinus: is it from NE Brazil or SE? According to Peters, Pinto
restricted the name to Bahia, so presumably that means this unique voice type
from NE Brazil is true murinus
(assuming Pinto did his homework!), leaving the more widespread
form, including the migratory populations Nacho talks about, as
ignobilis (which was
unhelpfully synonymized by HBW, clearly a move that must be reversed!), which
sounds more like wagae, but still
seems distinctive to my ear.
“The long and short of it is: this
complex is a mess, and I think this all needs to be ironed out before we go
splitting the murinus/incomtus
complex, even if it remains paraphyletic with respect to
N. ridgwayi... In some ways, this study mirrors that of
Nyctiprogne in having a
phylogeny that needs some "ground-truthing" to make sense of the
patterns it draws. So, with all that, I am changing my vote for Part 3 to NO.
We need so much more information to make that call!”
Additional comments from Robbins: “Yes, indeed this is a
mess that needs to be sorted out. Given what Dan has underscored, I’ll change
my 956.3 vote of splitting incomtus to a NO until things get clarified.”
Comments from Niels Krabbe (voting for Remsen); “As
pointed out by Dan, Mouse-colored Tyrannulet encompasses a larger number of
populations of different vocal types than the literature would suggest, and for
many of these there is still too little material available to properly define
them.
“After listening to recordings, I must agree that the call and
dawn song of maranonicus are strikingly different from those of tumbezana
occurring on the same slope, but it is unsatisfactory that this suggested split
is based partly on Lane's and Angulo's personal comments. So as a reminder that
SACC proposals should be based on published material, I cannot vote for this
split at present. That Nesotriccus has priority for the entire complex
is, as shown by both Zucker et al. (2016) and Harvey et al. (2020), indeed,
unquestionable.
“So my vote goes:
“Options 1-4: NO
“Option 5: YES apply the generic name Nesotriccus for the entire
complex.”
Comments from Glenn Seeholzer (voting for Jaramillo): “In
general I feel that the perfectly sampled, integrated taxonomic treatise should
not be the standard for every proposal. Very few have the resources or time to
indulge in such work meaning that obviously necessary and justified changes
will be stalled for lack of a peer-reviewed paper stating what was obvious at
the start. On the other hand, the split in Part 3 (already instituted by the
Clements/eBird taxonomy) feels hasty given what Dan and Shaun have uncovered so
this definitely feels like a case where more in-depth vocal analysis is
necessary.
“YES to Parts 1 and 2 - This split would be based primarily on the
assumption of close parapatry between the aligned clusters of vocal, plumage,
and genetic traits represented by tumbezana/inflava and maranonica.
A quick review of recordings on Xeno-canto and ML shows that the distinctive maranonica
vocal group is clearly present in the upper elevations of the west slope in
Piura (see Dan's great documentation here at 1900m) and published evidence that tumbezana
occurs to at least these elevations nearby (Angulo et al. 2011). While it would
be nice to have this better documented, I think the burden of proof is on those
who would say that these groups are not distinct enough, are not actually in
contact, or are somehow hybridizing and should be lumped.
“NO to Parts 3 and 4 - This
would create a paraphyletic incomta/murina with respect to ridgwayi,
which I'm fine with because paraphyly is compatible with the BSC. Ridgwayi
is clearly a diagnosable unit, but it is unclear what the taxonomic units are
among the constituent incomta + murina taxa. While the geographic
extremes of incomta/eremonoma vs. murina/wagae/ignobilis
may be quite different at their extremes, as Nacho states in the proposal,
Shaun and Dan seem to have uncovered more complex patterns of vocal variation
with potential evidence of intermediates (wagae) and multiple distinct
vocal groups in each putative species. For the same reasons that it seems
premature to split murina/wagae/ignobilis with so little
documentation (Part 4), it seems premature and potentially incorrect to split incomta/eremonoma
and murina/wagae/ignobilis (Part 3). I feel a detailed
published analysis of vocal variation is warranted in this case to define the
diagnostic vocal traits of the units and their geographic distributions before
considering further splitting.
“YES to part 5 - Nesotriccus has priority over Phaeomyias,
so ciao Phaeomyias and long runs of vowels.”