Proposal (967) to South
American Classification Committee
Note from Remsen: This is a proposal submitted to and passed unanimously
(10 to 0) by NACC (2023-B-2), modified by me slightly for SACC.
Transfer the subspecies minor and cinerascens from Myiodynastes chrysocephalus to M.
hemichrysus, thereby adding Myiodynastes
hemichrysus to SACC list
The NACC Checklist currently lists four species
of flycatchers in the genus Myiodynastes,
including two species, M. hemichrysus and
M. chrysocephalus, identified as
constituting a superspecies. These species are sometimes considered to be
conspecific (e.g., Cory and Hellmayr 1927), but when they are separated, M. hemichrysus of Costa Rica and western
Panama is generally treated as
monotypic, whereas M. chrysocephalus includes, in addition to nominate chrysocephalus of the Andes of Peru,
Bolivia, and northwestern Argentina, subspecies cinerascens of northern Colombia and Venezuela and subspecies minor of extreme eastern Panama south to
extreme northern Peru. Although the subspecies included in each species are not
specified in the accounts in AOU (1998), it is clear from the distributional
statements that this is the subspecies arrangement.
Plumage is somewhat variable within the complex.
According to Cory and Hellmayr, hemichrysus:
“differs immediately from the southern race by
the much deeper yellow, wholly unstreaked under parts, leaving only the chin
white, and by lacking the conspicuous rufous edges to the rectrices. In
wing-markings it more nearly resembles M.
c. chrysocephalus.”
The typical allocation of subspecies thus
follows the most conspicuous plumage features, and this allocation is followed
by most global sources (e.g., Dickinson and Christidis 2014, Clements et al.
2021, Gill et al. 2021). Birdlife, however, has recently partitioned the
subspecies in a different way, transferring minor
and cinerascens to M. hemichrysus. The rationale for this
change was as follows:
“Races minor and cinerascens here transferred from M. chrysocephalus because their dawn songs (repeated
“kweee!-tee-tu”) and daytime songs (strident “skeeeuw!”) are identical to those
of present species [i.e., M. hemichrysus]
and very different (scores of 3 for each) from those of chrysocephalus (respectively “kwee!-tlu-tee” and “ku-weet!..
weet!”, the “weet!” sharply rising and much higher-pitched than first note)…”
They further noted that M. chrysocephalus also differs from M. hemichrysus by its longer wing and tail and much buffier and
more restricted rufous edges to the tail.
The vocal descriptions above are condensed
versions of the conclusions of Boesman (2016), who compared vocalizations of
all four taxa in this complex. The similarity of the dawn and day songs of hemichrysus, minor, and cinerascens are
apparent from his figure and measurements comparing the vocalizations (see
following pages), as are the differences with chrysocephalus. Boesman’s summary of the differences is as follows:
“Voice
of Myiodynastes hemichrysus is about
identical to M. c. minor and M. c. cinerascens. We would need a large
number of samples to prove any consistent difference, but in any case it would
be very small. (Possibly the note shape is slightly different, with M. hemichrysus having a little notch at
the right side of the day-time song). Difference score for these taxa is thus 0
(or possibly 1).
“Difference
with chrysocephalus at the other hand
is quite noticeable: Day-time song has 2 (or 3) distinct notes (score 3) with
very different note shape (score 1) and slightly longer overall length (score
1). Dawn song ends with a fairly emphasized rising note (unlike all other races
which end in subdued notes) (score 2) and note shape of first note different
(score 1). The fact that both dawn song and day-time song are clearly different
makes this case even more convincing.”
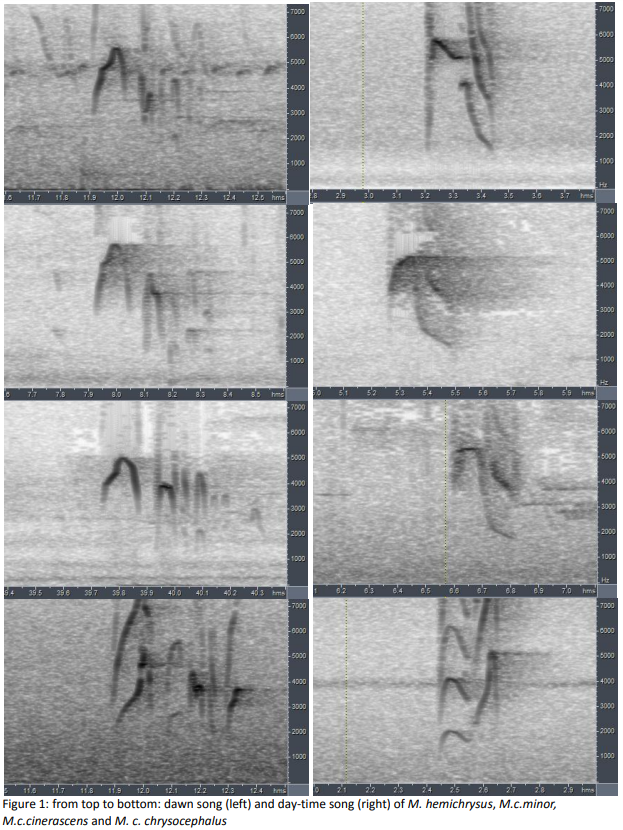
“Myiodynastes hemichrysus
dawn-song:
a repeated "kwee!-tee-t-tu" (n=1)
min.
freq. 2140Hz
max.
freq. 5600Hz
total
length 0.39s
length
1st note 0.13s
“day-time
song: a repeated loud strident "skeeew!" (n=6)
min.
freq. 1200-1550Hz
max.
freq. 5350-5830Hz
total
length 0.18-0.25s
“M. c.
minor
dawn-song:
a repeated "kwee!-tee-tu" or "kwee!-tee-tu-ti-lu" (n=3)
min.
freq. 1380-1590Hz
max.
freq. 5290-5740Hz
total
length 0.32-0.43s
length
1st note 0.12-0.14s
“day-time
song: a loud strident "skeeeuw!" (n=6)
min.
freq. 1030-1450Hz
max.
freq. 5000-5550Hz
total
length 0.15-0.29s
“M. c.
cinerascens
dawn-song:
a repeated "kwee!-tee-tu" (n=2)
min.
freq. 1300-1320Hz
max.
freq. 5030-5120Hz
total
length 0.37-0.39s
length
1st note 0.15-0.16s
“day-time
song: a loud strident "skeeew!" or "skeeeuuw!" (n=6)
min.
freq. 1060-1400Hz
max.
freq. 4540-6340Hz
total
length 0.17-0.28s
“M. c.
chrysocephalus
dawn-song:
a repeated "kwee!-tlu-tee" (n=2)
min.
freq. 1950-2050Hz
max.
freq. 5220-5300Hz
total
length 0.42-0.46Hz
length
1st note 0.12-0.13s
“day-time
song: a loud strident "ku-weet!" or "ku-weet!.. weet!"
(weet! sharply rising and much higher-pitched than first note) (n=8)
min.
freq. 1150-1380Hz
max.
freq. 4000-5300Hz
total
length 0.22-0.66s
# of
notes 2-3”
Sample sizes for
these comparisons aren’t huge, especially for dawn songs, but recordings
available on the xeno-canto and Macaulay Library websites confirm the
differences between the two species and clearly place minor and cinerascens with
M. hemichrysus rather than M. chrysocephalus.
These differences were also recognized in the
field guide to the birds of Peru (Schulenberg et al. 2007), in which a clear
distinction was made between the vocalizations of minor of northern Peru and those of the more widespread chrysocephalus, although without
comparing the vocalizations of minor with
those of Central American hemichrysus.
Recommendation:
I recommend that we transfer subspecies minor (and extralimital subspecies cinerascens) from M. chrysocephalus to M.
hemichrysus. Although plumage favors the traditional placement of minor and cinerascens in chrysocephalus,
this is contradicted by the vocal data, which indicate that these two
subspecies belong in M. hemichrysus.
Vocalizations are innate and primary indicators of species limits and
affinities in suboscine birds, and in this case they demonstrate that the
traditional allocation of subspecies in the complex was in error. If adopted,
this change would add M. hemichrysus to
the SACC main list.
References:
Boesman, P. 2016. Notes on the vocalizations of Golden-crowned
Flycatcher (Myiodynastes chrysocephalus)
and Golden-bellied Flycatcher (Myiodynastes
hemichrysus). HBW Alive Ornithological Note 141. In: Handbook of the Birds
of the World Alive. Lynx Edicions, Barcelona. (retrieved from http://www.hbw.com/node/932065 on 10 August 2016).
Schulenberg, T. S., D. F. Stotz, D. F. Lane, L. B. McQueen, J. P.
O’Neill, and N. J. Schmitt. 2007. Birds of Peru. Princeton University Press,
Princeton.
Terry Chesser, February 2023
Comments from Remsen (as in my NACC comments):
“YES. Of course we would prefer a peer-reviewed paper
on those vocalizations, but I think the differences shown by Boesman, and
consistent with those mentioned by Schulenberg et al., shift the burden of
proof to justifying maintaining the current treatment. Voice is everything in cases of species
limits in parapatric and sympatric tyrannids, and so I think we are on solid
ground to revise our classification based on the latest information on voice.”
Comments from Robbins: “YES.
Both dawn song and calls support transfer of the subspecies of minor and
cinerascens from Myiodynastes chrysocephalus to M. hemichrysus.
I agree we don't need to wait for a peer-reviewed paper to make this move.
Needless to say, it will be interesting to eventually see what genetic data
demonstrate.”
Comments from Lane: “YES to
shifting minor and cinerascens to M. hemichrysus,
leaving M. chrysocephalus monotypic. The voice here makes this a
fairly easy decision, but I do chafe on one detail: some verbiage has been
spent discussing dawn song distinctions, but I cannot locate a recording of
dawn song from true hemichrysus (which seems surprisingly poorly sampled
in general), and Boesman's voice assessment
maddeningly does not specify what recordings he reviewed (including which are
the sources of the sonograms)! I really would like to compare the dawn songs of
hemichrysus and minor just to feel a little better about this
taxonomic shift.”
Comments
from Areta: “YES to a monotypic M.
chrysocephalus and to
moving minor
and cinerascens
to M.
hemichrysus, the differences between the northern and southern South American
populations of "chrysocephalus" have puzzled me for several
years, and were in my "to do" list. I never compared songs against
hemichrysus, but the decision is straightforward. As per
the Boesman pieces, I agree with Dan, and I have also repeatedly pointed out
their deficiencies, which is a pity, given the amount of work performed.”
Comment from Peter Boesman: “I noticed in the proposal
967 (Golden-bellied Flycatcher complex) that Dan Lane commented he could not
find a dawn song of the hemichrysus
taxon. There is one in
Macaulay Library:”
ML144068
Golden-bellied Flycatcher Macaulay Library
Comment
from Stiles: “YES
to moving these two subspecies from chrysocephalus into hemichrysus:
in such a case, vocalizations clearly trump the rather weak plumage
differences.”
Comment
from Del-Rio (who has Pacheco vote): “YES.”
Comments
from Claramunt: “NO. Plumage is also innate and primary indicator of species limits
and affinities in suboscine birds. If there is a conflict between vocal and
plumage data, then we should examine that conflict and seek additional data to
clarify the situation, not accept whatever the vocal data suggest at face
value.”
Comments
from Bonaccorso: “NO. We need to aim toward a more integrative taxonomy when making
these decisions. Plumages tell one story, songs tell another, and we don´t have
genetic or other data to solve the ‘tie.’”
Comments
from Oscar Johnson (voting for Jaramillo): “YES, with a nod toward
the concerns raised by others about the discordance between plumage and song.
Both plumage and song do of course have a genetic basis in suboscines, but
given how important song is for mate recognition, I'm much more inclined to place
emphasis on song in species delimitation for species complexes in which plumage
is broadly similar. It is hard to argue with the similarity in vocalizations,
which are really nearly identical in all taxa north of the Marañón Valley, and
quite distinct to the south. The song differences in particular are remarkably
consistent, but I will note that there does appear to be some variation in the
daytime calls, with some recordings of hemichrysus/minor/cinerascens having some rising notes, but these are usually
given singly and not in combination with the arched note as in
chrysocephalus. That said, I agree that nominate
hemichrysus is quite distinctive in plumage compared to all
other taxa in the group, and I wonder about the possibility of a three species
treatment; hemichrysus, minor (including
cinerascens), and chrysocephalus, which would emphasize the distinct combinations of
differences in both song and plumage. More work is clearly needed.”